


A review on decellularization scaffolding for liver regeneration in transplantation
 0
0 Abstract
Chronic liver disease constitutes a major global health burden, driven largely by complications of cirrhosis, viral hepatitis and hepatocellular carcinoma. While liver transplantation remains the current definitive treatment, its impact is constrained by organ scarcity, strict eligibility criteria, and lifelong immune-related complications. Through comparison of transplant strategies and decellularization-recellularization protocols, this review evaluates the translational potential of decellularized liver scaffolds, with particular emphasis on their feasibility and reported outcomes in surgical transplantation. Decellularization approaches varied across studies, with differences in preferred cell sources and recellularization outcomes. Thrombogenicity and biliary system reconstruction remain key challenges requiring further investigation. Perfusion-based recellularization in bioreactor systems, provides controlled cell seeding and dynamic culture conditions. In this context, segmental or split liver scaffolds may serve as a viable model for future translational studies. Overall, decellularized liver scaffolds represent a promising bridging strategy to address the current limitations. However, there is heterogeneity between protocols and important translational barriers remain. Further research is required to define the optimal methods through validation in large-animal transplantation studies.
Keywords
INTRODUCTION
Chronic liver disease is estimated to be responsible for more than two million deaths globally per annum[1]. Around half of this disease burden occurs in the Asia-Pacific region[2].
As chronic liver disease progresses and end-stage liver disease ensues, the range of current treatment options is limited largely to liver transplantation. The clinical utility of organ transplantation is limited by donor organ availability, the need for lifelong immunosuppression, and the exclusion of certain patients with hepatocellular carcinoma[3].
These constraints have fuelled research into novel organ replacement strategies. In recent years, decellularized liver scaffolds have emerged as the most promising approach. The process of developing decellularized liver scaffolds involves the removal of cellular material from the tissue, using extraction techniques that preserve the native extracellular matrix (ECM) architecture and biochemical composition[4]. Current liver scaffolds are based on either xenogeneic liver (porcine or rodent) or human donor liver considered unsuitable for transplantation[4]. Once the scaffold has been decellularized, it is recellularized with human cellular material and can support sustained cell survival[5-7]. This review summarises the current state of liver decellularization and recellularization techniques.
ORGAN SHORTAGE AND LIMITS OF CURRENT TRANSPLANTATION APPROACHES
Liver transplantation remains the mainstay treatment for end-stage liver disease. However, it is constrained by organ supply. The currently applied techniques for liver replacement therapy comprise whole-liver transplantation, split-liver transplantation, and xenotransplantation. Each is considered here.
Whole liver transplant
Whole liver transplantation has been considered the gold standard operative technique globally. Deceased-donor transplantation has been the principal organ source in North America and Western Europe. Living-donor transplantation has become popularised in Asian countries[3]. In an attempt to overcome organ shortage, the criteria for organ suitability have been relaxed, to permit inclusion of organs previously considered unsuitable (e.g., presence of steatosis, older age, viraemic donors, donation after circulatory death)[3,8,9]. Complications of whole organ transplantation include allograft dysfunction, biliary complications, and the morbidity due to lifelong immunosuppression[10-12].
Split-liver transplantation
Split-liver transplantation (SLT) involves dividing a single deceased-donor liver into two partial grafts, allowing for greater organ availability[13]. This approach has proven particularly suited to pediatric recipients and selected adult patients[14,15]. SLT is considered technically more challenging than whole liver transplantation[8,14]. Higher rates of biliary and vascular complications have been observed with SLT[14,16].
Machine perfusion
Ex vivo liver perfusion (EVLP) is an emerging preservation approach in liver transplantation, in which the graft is perfused via an extracorporeal circuit through the hepatic vasculature. This technique has been shown to reduce ischemia-reperfusion injury. It also allows viability testing of the organ prior to transplantation. The two approaches used for EVLP are normothermic machine perfusion (NMP) and hypothermic oxygenated perfusion (HOPE)[17,18]. Both have been shown to improve metabolic function of the graft and extend organ preservation time before surgical transplantation[19-21].
Emerging research: xenotransplantation
Xenotransplantation involves the use of organs from other species, predominantly genetically engineered pigs. Advances in gene editing targeting glycan xenoantigens (alpha-Gal and other non-Gal xenoantigens) have reduced the risk of hyperacute rejection, while immunosuppressive strategies such as costimulation blockade have further improved short-term xenograft compatibility and survival[22,23]. However, the immunogenicity of these organs remains the limitation to this approach currently[24,25]. Immunologically mediated coagulation-related complications, such as thrombotic microangiopathy, can result in graft failure[26,27]. Other risks of xenotransplantation include the theoretical risk of zoonotic transmission, and underproduction of bile and porcine albumin for sustained functional support[25,26]. Although potentially promising as a future adjunct, xenotransplantation remains experimental at present.
These inherent limitations of organ transplantation have promoted research into liver scaffolds as suitable alternatives.
CORE PRINCIPLES OF LIVER DECELLULARIZATION
The principles underlying liver scaffolds are creation of a non-immunogenic, three-dimensional scaffold with preservation of vascular and biliary anatomy. This decellularized tissue is then repopulated with new cell types resulting in an engineered construct that mimics the native original tissue[28].
Organ source selection
A number of different source tissues have been used in liver decellularization over the last decade. These have included rodent tissue, porcine liver and discarded human liver tissue from ineligible donors[29,30]. Porcine liver has been used largely because of its size and anatomical compatibility with human liver. As alluded to previously, a pitfall of xenogeneic scaffolds is their immunogenicity due to xeno-epitopes not found in humans. Certain epitopes have proved easier to remove than others (such as the alpha-gal epitope)[7].
In contrast, discarded human liver-derived ECM may offer lower immunogenicity[31]. It is derived from a more heterogenous population, where age and disease-related ECM remodeling (e.g., fibrosis), can affect its resistance to enzymatic processing of the ECM[32].
Decellularization agents and protocol
Decellularization commonly involves a combination of physical, chemical, and enzymatic approaches. The aim is to disrupt cell membranes, induce cell death and facilitate removal of cellular material[33]. It is a delicate balance between the use of aggressive agents that achieve a more complete decellularization process, and the resultant damage to key ECM components and vascular architecture[7].
Chemical methods
Detergents: sodium dodecyl sulfate (SDS) and Triton X-100
Chemical decellularization relies on detergents to solubilize cell membranes by the disruption of lipid-lipid interactions. The most commonly used agents are non-ionic detergents (Triton X-100) and ionic detergents (SDS). Triton X-100 is considered less aggressive because it disrupts mainly lipid-lipid and lipid-protein interactions while preserving protein-protein interactions[4]. SDS is considered more aggressive in solubilizing cellular membranes and denaturing proteins[4]. Thus, SDS achieves more robust removal of cytoplasmic and nuclear remnants, at the risk of compromise of ECM integrity. In a study comparing their effect on the liver scaffold, Willemse et al. found that Triton X-100-only perfusion allowed preservation of 1.5× more collagen and 2.5× more sGAG (Glycosaminoglycans) than a combination Triton X-100 and SDS perfusion[34]. However, non-ionic detergents are not completely ECM-sparing. A study by Jiang et al. noted that despite apparently preserved collagen networks, Triton X-100 can still deplete key ECM constituents[35]. These findings suggest that the balance between SDS and Triton X-100 may be an important determinant of decellularization outcomes.
Acids and alkalis
These chemical reagents act by solubilizing cytoplasmic components facilitating the removal of nucleic material. The most studied among these include peracetic acid (PAA) which has been frequently used as an adjunct to primary detergent-based protocols[6,36]. Of alkaline reagents studied, NaOH has proven the most efficient at extracting cellular components such as chromosomal and plasmid DNA. Its greater decellularization properties come at the expense of greater ECM damage[36-38].
Hypotonic/hypertonic solutions
Hypotonic/hypertonic decellularization alters low and high osmolality to induce osmotic shock through cell swelling and shrinkage, while maintaining comparatively limited perturbation to the ECM architecture[37,39]. Their decellularization properties are less effective than other methods; however, it is likely that osmotic shock will be used as an adjunct to other methods rather than as a stand-alone approach for complex and thicker tissues[37,40].
Others
Aside from these classical detergents, recent protocols have reported use of chelates, e.g., ethylenediaminetetraacetic acid (EDTA), alcohol and saponin as potential adjunct agents for decellularization[4,41,42].
Physical methods
The physical decellularization approach involves the use of membrane rupture by energy input e.g., thermal, pressure or mechanical, followed by washing to remove debris. Freeze-thaw cycling is the most established pre-step. It creates rapid thermal change and ice crystals to induce cell membrane lysis. Many current protocols start with at least one freeze-thaw cycle prior to use of chemical agents[43,44]. Other putative decellularization techniques include use of ultrasound and electroporation[7,44,45].
Enzymatic methods
Enzymatic decellularization is typically used as an adjunct to the previously described physical and chemical agents. The most commonly applied enzymes are nucleases and trypsin. Nucleases degrade residual nucleic acids and reduce the potential immunogenicity of the scaffold[4]. Indeed, recent studies by Ijima et al. and Stefan et al. have included deoxyribonuclease (DNase)/ribonuclease (RNase) treatments after Triton X-100-based protocols. These have reported satisfactory decellularization with good preservation of vascular architecture and key ECM features[46,47]. For protein hydrolysis, trypsin is typically used to accelerate cell detachment from adhering tissue. A recent example by Faccioli et al. showed that trypsin used in combination with other detergents enabled efficient cell removal and preservation of ECM components in porcine liver[48]; however, as trypsin is potentially disruptive to ECM, optimisation of concentration and exposure is critical to avoid excessive damage[7].
RECELLULARIZATION
Recellularization is the process whereby the decellularized scaffold is reconstituted with specific cell types to form a functional construct that mimics the function of the original tissue[7]. A number of different cell sources have been used for recellularization. These are summarised in Table 1, with the merits and demerits of each shown.
Comparison of cell sources utilized for whole decellularized liver recellularization
| Cell source | Advantages | Limitations | Translational readiness level | References |
| Primary human hepatocytes | Gold standard for functionality (albumin and urea production, ammonia clearance) High host compatibility | Scarce availability Poor proliferative capacity in vitro Rapid phenotypic dedifferentiation and loss of specific functions during culture | Preclinical (Large animal models) Large animal (porcine) heterotopic bioengineered liver transplants Limited by donor shortage | Hussein et al., 2024[7] Anderson et al., 2021[49] Higashi et al., 2022[50] |
| HUVECs | Accessible with non-invasive harvest from medical waste with high isolation yield Low risk in tumorigenicity Support re-endothelialization, maintain vascular patency, effectively reduce platelet adhesion and initial thrombosis | Donor variability Difficult to obtain autologous cell source Allogenic/Xenogenic recipients may present with strong rejection Poor long-term engraftment | Preclinical (Large animal models) Widely utilized for re-endothelialization in rodent and porcine liver scaffolds Further evaluated in ex vivo blood perfusion and large-animal in vivo transplantation | Shaheen et al., 2020[5] Hussein et al., 2024[7] Toprakhisar et al., 2023[29] Anderson et al., 2021[49] Horie et al., 2025[51] |
| Human fetal hepatocytes | High proliferative capacity and rapid engraftment Low risk in tumorigenicity | Incomplete functional maturation Ethical concerns Limited availability | Preclinical (In vitro/Small animal models) Restricted to preclinical studies due to severe ethical constraints and limited cell supply | Hussein et al., 2024[7] Liu et al., 2024[52] Afzal et al., 2023[53] |
| Immortalized and Hepatoma cell lines (e.g., HepG2) | Highly available, rapid growth, and easy to expand. Homogeneous population with phenotypic stability | Tumorigenic origin restricts its clinical translation May have suboptimal metabolic function | Preclinical (in vitro only) Restricted to in vitro drug screening, toxicity testing, and proof-of-concept recellularization | Hussein et al., 2024[7] Toprakhisar et al., 2023[29] Liu et al., 2024[52] Afzal et al., 2023[53] Afrin et al., 2025[54] |
| Mesenchymal stem cells | Hepatocyte-like differentiation potential Immunomodulatory Observable cell infiltration | Limited mature hepatic function Restricted engraftment | Preclinical use in recellularized whole organs | Toprakhisar et al., 2023[29] Afzal et al., 2023[53] Panahi et al., 2022[55] Kang et al., 2020[56] |
| iPSCs | Can be patient-specific (autologous), may reduce immune rejection Pluripotent potential to differentiate into multiple liver lineages (hepatocytes, cholangiocytes, endothelial cells) | Functional immaturity (often arrest at a fetal phenotype) Bile canaliculi reconstruction unconfirmed Inherent risk of tumorigenicity and teratoma formation | Preclinical (Small animal models) Demonstrated short-term in vivo function in rodent auxiliary transplant models Requires extensive validation before human clinical trials due to tumorigenic risks | Hussein et al., 2024[7] Toprakhisar et al., 2023[29] Takeishi et al., 2020[57] Acun et al., 2022[58] |
Those showing the most promise have included primary human hepatocytes, which exhibit a high level of host compatibility. At a clinically relevant scale, Anderson et al. re-endothelialized porcine whole-liver scaffolds with primary hepatocytes with or without human umbilical vein endothelial cells (HUVECs)[49]. Under perfusion culture, the resulting constructs exhibited hepatic function, including albumin and urea secretion and ammonia clearance. Higashi et al. using engineered porcine-derived scaffolds with primary hepatocytes have reported functional benefit following auxiliary transplant in an induced liver failure pig for 28 days[50].
Human fetal hepatocytes or hepatic endothelial cells hold appeal because of their low risk of tumorigenicity and zoonotic transmission[7]. Liu et al. seeded decellularized rat liver scaffolds with human fetal liver cells from elective terminations of pregnancy and reached measurable drug metabolic capabilities and enhanced cellular organization within 7-14 days[52].
Mesenchymal stem cells (MSC) are known for their self-renewal capacity and immunomodulatory properties[56]. They can be obtained relatively easily from sources such as bone marrow, umbilical cord, and adipose tissue[59]. Preclinical evidence indicates that MSC-to-hepatocyte differentiation accounts for only a minor fraction of overall liver mass, limiting their capacity to function as an independent repopulating cell source[56]. In the rat scaffold study (Kadota et al.), co-infusion of hepatocytes with bone marrow-derived MSCs was reported to enrich the regenerative microenvironment[7,60].
Induced pluripotent stem cells (iPSCs) are an emerging cell source for consideration in recellularization strategies as they can differentiate into hepatic cell lineages. Takeishi et al. repopulated decellularized rat liver scaffolds with iPSC hepatocytes, endothelial and biliary cells. The authors achieved short-term in vivo function after auxiliary transplantation in immunocompromised rats[57].
Reconstruction of the biliary tree has proved to be challenging in the creation of a functional bioengineered liver graft. This process requires repopulation of the decellularized scaffold with cholangiocytes to preserve normal hepatic physiology and bile transport to minimise post-transplantation injury. Chen et al. perfused decellularized rat liver scaffolds with primary cholangiocytes to develop functional ductal organoids[61]. The reconstructed biliary cells remained viable and expressed enriched biomarkers. Bile secretion and transport functions were established by day 14.
EX VIVO PERFUSION AND CURRENT STATUS OF TRANSPLANTATION STUDIES
In liver bioengineering, recellularization may be achieved by one of two methods, either direct injection into the parenchyma or by infusion into the vascular system[7]. With its unique portal circulation, perfusion-based delivery is well-suited to achieve widespread cell distribution. Ex vivo machine perfusion is the process by which the liver scaffold is perfused through an extracorporeal circulation. Similarly, perfusion bioreactors support oxygen and nutrient delivery through preserved vascular conduits under controlled perfusion pressure, while providing an ex vivo environment for graft maturation and pre-implantation functional assessment[53].
Application of ex vivo perfusion on decellularized liver scaffolds on rodent and porcine models
Table 2 summarises the most recent studies that employ perfusion recellularization in rodent and porcine liver models. In some instances, the experimental liver constructs underwent ex vivo vascular patency testing and transplantation. This approach allows comparisons to be made as to how perfusion recellularization protocols relate to in vivo liver function.
Comparative summary of perfusion recellularization studies assessing vascular patency via ex vivo blood perfusion followed by transplantation
| References | Model | Recellularization method | Ex vivo hemocompatibility assessment | Transplantation | Outcomes |
| Shaheen et al., 2020[5] | Whole-organ porcine | HUVECs endothelial cells were seeded via the SVC, followed by portal vein after 24 h Graft was then cultured and continuously perfused in bioreactor at 12 mmHg up to 30 days | Ex vivo blood loop circuit with fresh heparinized porcine blood was used to assess vascular patency | Heterotopic auxiliary transplant in immunosuppressed pig recipients | Re-endothelialized grafts maintained perfusion for over two weeks Peak glucose consumption rate > 30 mg/h during bioreactor culture Endothelial cells populated sinusoids and expressed markers similar to normal liver tissue |
| Anderson et al., 2021[49] | Whole-organ porcine | HUVECs endothelial cells were seeded via sIVC, followed by PV after 24 h Porcine hepatocytes infused via the bile duct 13-16 days later in a perfusion bioreactor | Ex vivo perfusion circuit with fresh heparinized porcine blood was used to assess vascular patency | Heterotopic auxiliary transplant in a surgically induced pig model with acute liver failure | Continuous perfusion was achieved with implantation duration of 48 h Scaffolds are competent in supporting hepatocyte function, including albumin and urea production and ammonia detoxification |
| Sassi et al., 2021[62] | Whole-organ rat liver scaffold | HepG2 and primary human hepatocytes were seeded via PV, and cultured in a novel perfusion-based bioreactor | - | - | Perfusion reactor improved overall cell viability and function Capable of supporting cell survival and metabolism |
| Higashi et al., 2022[50] | Whole-organ porcine | Primary hepatocytes were seeded via IVC Endothelial cells were seeded via PV Both cells used closed-chamber perfusion with pressure and flow monitoring Graft maintained under extracorporeal perfusion for 4 days post-seeding | - | Heterotopic auxiliary transplant in pigs with induced liver failure | Bioengineered liver graft improved liver function for 28 days and exhibited upregulation of liver-specific genes Demonstrated 28-day post-transplant function of an engineered liver graft in a large-animal model, supporting its clinical translational potential |
| Nelson et al., 2023[63] | Whole-organ Porcine | HUVEC endothelialization via the sIVC and PV in a perfusion bioreactor for 8-16 days, followed by primary human or porcine liver cell seeding with continued perfusion culture | Ex vivo perfusion circuit with fresh heparinized porcine whole blood was perfused via the PV for 180 min at 37 °C with an oxygenator | - | Grafts seeded with primary human liver cells showed lower perfusion pressures and greater ammonia clearance than porcine-cell grafts during blood perfusion testing |
| Campinoti et al., 2023[64] | Whole-organ rat liver scaffold | Human AEC seeded via the PV and cultured in a perfusion reactor | Whole liver vasculature was flushed with PBS then gradually perfused with 1% trypan blue | - | AEC-derived cells expressed hepatic markers and showed functional hepatocyte activity Perfusion bioreactor culture supported hepatocyte differentiation through sustained oxygenation and media perfusion |
| Liu et al., 2024[52] | Whole-organ rat liver scaffold | Human fetal liver cells and endothelial cells co-seeded via the PV, followed by perfusion culture and maturation for 36 h | - | - | Recellularized liver showed enhanced cellular organisation and metabolic function, supporting their potential use for drug metabolism and toxicity studies |
| Yadav et al., 2024[65] | Whole-organ rat liver scaffold with heparin immobilization | EA.hy926 endothelial cells were seeded via PV and cultured in a bioreactor for 7 days | Ex vivo blood perfusion and platelet adhesion studies | - | Heparinization reduced thrombogenicity and platelet aggregation, enhanced endothelial adhesion and promoted angiogenesis |
| Afrin et al., 2025[54] | Whole-organ rat liver scaffold with perfusion-assisted fibronectin coating | EA.hy926 endothelial cells and hepatocellular carcinoma (HepG2) cells were seeded via PV and cultured in a bioreactor for 7 days | Ex vivo whole blood perfusion with clot assessment, histology, and platelet adhesion assays for thrombogenicity assessment | - | Perfusion assisted fibronectin coating technique improved: endothelial cell adhesion within blood vessels, reduced thrombogenicity, and increased HepG2 cell proliferation |
| Horie et al., 2025[51] | Whole-organ rat liver scaffold modified with biomimetic polymer - MPC | HUVEC re-endothelialization via PV perfusion, followed by perfusion culture | Ex vivo blood perfusion for platelet deposition | Heterotopic transplantation | MPC treatment reduced platelet deposition and thrombus formation during blood perfusion and in the heterotopic transplant model |
| Guo et al., 2023[66] | Whole-organ rat liver scaffold | HUVEC implanted via PV hBMSCs and mouse hepatocyte cell line implanted via the bile duct Scaffolds were perfusion-cultured in a bioreactor at a rate of 1 mL/min | Endothelial barrier integrity assessed via blood leakage test | Orthotopic transplantation | Functional bioengineered livers showed biosynthesis and metabolic activity Implanted hBMSCs and hepatocyte cell lines demonstrated good alignment with the parenchyma Orthotopic transplantation achieved a longer survival time compared to the control group |
Thrombogenicity mitigation strategies for decellularized liver scaffolds
Mitigation of the risk of organ thrombosis remains a key requirement for the translational feasibility of decellularized liver scaffolds.
Re-endothelialization represents an important strategy for improving vascular patency and reducing scaffold thrombogenicity. It allows restoration of the vascular lining and limits direct coagulation contact with the exposed ECM[7]. This approach has been explored in large-animal models with encouraging translational progress. Shaheen et al. seeded decellularized liver scaffolds with HUVECs and achieved sustained perfusion in vivo after heterotopic transplantation for up to 15 days[5]. Similarly, Higashi et al. repopulated the vascular compartment of a porcine liver scaffold with endothelial cells, and reported extended survival of the transplanted liver graft up to 28 days[50]. Liver sinusoidal endothelial cells (LSECs) may also be ideal for reconstructing the sinusoidal vascular network, however large-scale applications have not yet been established[50]. These findings suggest endothelialization as a promising method to improve revascularization, however further optimisation of endothelial cell source and long-term vascular stability is still required.
Heparin has been used as an anti-thrombotic agent. Yadav et al. found it to reduce thrombogenicity and platelet aggregation during ex vivo blood perfusion, while also improving endothelial adhesion[65]. Fibronectin coating has been explored as another potential anti-thrombotic agent. Afrin et al. found that fibronectin coating on the liver scaffolds resulted in improved endothelial cell adhesion within the vascular network, reduced thrombogenicity, and increased HepG2 proliferation[54]. This model has not been trialled in a xenotransplant model, so its in vivo performance remains uncertain. In a different approach, Horie et al. applied the biomimetic polymer 2-methacryloyloxyethyl phosphorylcholine (MPC) to blood-contacting ECM surfaces and reported reduced platelet deposition and suppressed thrombus formation during ex vivo blood perfusion, with similar antithrombotic effects maintained in a heterotopic transplantation model[51].
Collectively, these studies suggest potential approaches to targeting scaffold thrombogenicity. However, longer term hemocompatibility testing in large-animal transplantation models remains necessary before clinical translation can be considered.
TRANSLATIONAL BARRIERS TO THE CLINICAL TRANSPLANTATION OF DECELLULARIZED AND RECELLULARIZED LIVER GRAFTS
Although the reported decellularization and recellularization technologies employed for bioengineered liver grafts have demonstrated proof-of-concept potential in vitro and in animal models, translation of this work into whole-organ bioengineering remains a challenge. Potential barriers to the introduction of this new technology include the following: in vitro, intraoperative, in vivo, regulatory, and economic.
In vitro barriers: producing functional liver grafts
Successful liver transplantation fundamentally depends on a well-functioning graft. However, current bioengineered liver constructs remain unable to consistently produce fully functional grafts. It is primarily hindered by the number of cells required and the profound difficulty in maintaining their functional maturity. To achieve a clinically relevant liver mass for an adult human liver, a cell amount of approximately 150 to 350 billion cells is required[30]. For a 70-kg patient, it requires at least 84 billion functional hepatocytes to achieve the 20%-30% hepatic mass necessary to sustain baseline physiological functions[7]. Although primary hepatocytes are the gold standard in most studies, their limited donor availability constrains clinical application. Moreover, primary hepatocytes still face donor-recipient immunological mismatch. In addition to these limitations, primary hepatocytes undergo rapid dedifferentiation ex vivo, with downregulation of metabolic enzyme synthesis reported within 24 h of culture[62].
To overcome the shortage of primary hepatocytes, patient specific iPSCs have been trialled. However, iPSC-derived hepatocytes frequently arrest development at a fetal phenotype and demonstrate incomplete functional maturation in critical parameters like cytochrome P450 activity[7,57]. Furthermore, the lack of an intact and functional biliary tree in most models leads to the accumulation of cytotoxic bile, hindering long-term survival[7,57].
As a result, current bioengineered liver grafts suffer from short in vivo survival times. Early preclinical models failed to maintain continuous perfusion for more than 3 days[5]. Although recent advancements utilizing dynamic bioreactors and immunosuppressed large animal models have extended the graft survival to 15 days and even 28 days, the overall hepatic function remains partial, and delayed graft failure due to cellular senescence and vascular stenosis remains unresolved[6,50].
Intraoperative barriers: surgical feasibility and hemodynamic stability
Although many small animal studies of bioengineered liver transplantation have demonstrated short-term graft survival and partial liver function, the clinical translation to human models requires validation in large animal models, particularly porcine models[6,67]. Several surgical technical problems remain in the transplantation of bioengineered livers. Firstly, the vessels of the decellularized liver scaffold are usually more fragile than those of native liver tissue. They are easily torn during vascular clamping, traction, or suturing, which makes vascular anastomosis more difficult.
Further, the liver scaffold must be able to immediately withstand physiological portal and arterial pressure after implantation. Many current large animal models have relied on auxiliary rather than total orthotopic transplantation, largely because the current bioengineered livers do not provide sufficient liver function to support complete liver replacement.
Once the graft is re-perfused with blood, exposed vascular ECM may rapidly activate the coagulation cascade and platelet aggregation, leading to acute thrombosis within minutes if the graft is not adequately re-endothelialized[6,49]. Even with several re-endothelialization strategies, achieving complete and uniform coverage of the complex sinusoidal microvascular network remains difficult, causing the graft to face a hostile environment of increased vascular resistance, incomplete microcirculation, and eventual ischemic failure[7,49].
In vivo barriers: dual-source immunogenicity and immune rejection
Even if surgical anastomosis is successful and acute thrombosis is avoided, the bioengineered graft faces severe immune responses derived from a “dual-source immunogenicity”: the decellularized scaffold and the repopulating cells.
From the scaffold perspective, xenogeneic matrices (e.g., porcine livers) may retain highly immunogenic carbohydrate epitopes, notably galactose-alpha-1,3-galactose (α-Gal), N-glycolylneuraminic acid (Neu5Gc), and Sda antigens[36]. Furthermore, structural damage to the ECM during decellularization releases damage-associated molecular patterns (DAMPs), such as low molecular weight hyaluronic acid and fibronectin fragments, which strongly activate pattern recognition receptors (PRRs) on host macrophages, driving them toward a pro-inflammatory (M1) phenotype[36].
From the cellular perspective, if the cells utilized for recellularization were not recipient-specific cells, the adaptive immune rejection may be provoked. Preclinical studies have shown that repopulating porcine scaffolds with human cells, such as HUVECs, triggers intense complement-mediated cytotoxicity and robust antibody responses. Without immunosuppression, the humanized grafts are rapidly destroyed within three to seven days post-transplantation[5]. Theoretically, recellularization with recipient-specific cells may reduce or even eliminate the need for immunosuppressive therapy[6]. In reality, however, even an autologous-derived cell source should not be considered entirely immune-privileged. MSCs are generally considered to have relatively low immunogenicity, yet they are not completely immunologically inert[68,69]. Similarly, autologous iPSC-derived cells may still elicit immune responses[70,71].
Regulatory barriers: insufficient regulations for whole organ engineered products
Regulations are established to protect public health and patient safety by strictly controlling the quality, safety, and efficacy of medical products. However, when emerging products or clinical scenarios fall outside the existing regulations, the researchers and the industry developers may lack clear guidance on study design, manufacturing processes, and defining appropriate quality-control and translational strategies[72]. The whole-organ bioengineered liver produced via decellularization and recellularization techniques is a complex product containing decellularized scaffold, implanted cells, and vascular structures sourced from either humans or animals. It also requires ex vivo processing to produce a functional graft. These features make whole-organ bioengineered liver grafts fundamentally more complex than conventional cell therapies or scaffold-based constructs[73].
In the United States, there is no specific regulatory framework established for whole organ engineering. For the bioengineered liver composed of human-source scaffolds and cells, the regulation may fall within the Food and Drug Administration (FDA) framework of human cells, tissues, and cellular and tissue-based products (HCT/P)[7,72,74]. Such products would exceed section 361 of the Public Health Service Act (PHS Act)[75]. In contrast, for animal based decellularized matrices seeded with human cells, the regulatory pathways remain unclear. Decellularized and recellularized bioengineered organs would more likely fall under section 351 of the PHS Act and/or the Federal Food, Drug, and Cosmetic Act (FD&C Act) as drug, device, and/or biological products[75]. They would be subject to stricter regulatory pathways, including current good tissue practice (cGTP), current good manufacturing practice (cGMP), and would generally require an Investigational New Drug (IND) application before clinical investigation in humans[72,75].
In the European Union, the bioengineered liver would most likely fit within the advanced therapy medicinal product (ATMP) framework, particularly as a tissue-engineered product (TEP), according to Regulation (EC) No 1394/2007[72,73,76,77]. Under Regulation (EC) No 1394/2007, a Committee for Advanced Therapies (CAT) within the European Medicines Agency (EMA) was established to prepare a draft opinion on the quality, safety and efficacy of each ATMP. Although the ATMP framework serves as the closest regulatory pathway for bioengineered products, the organ-level standards remain undeveloped. Whole-organ bioengineering products would still require case-by-case evaluation[77].
In Japan, two acts for human regenerative medicine were enacted in 2014: the Act on the Safety of Regenerative Medicine (ASRM) and the Pharmaceuticals, Medical Devices, and Other Therapeutic Products Act (PMD Act)[72,78,79]. The ASRM mainly regulated the clinical use and medical research for regenerative medicine in clinical institutions, while the PMD Act is more related to the development and commercialization of cell-containing bioengineered products[80]. This dual-track system in Japan appears relatively favourable in regenerative medicine research and product development compared with other regions. Nevertheless, both acts were originally designed for cell-based therapy, rather than for whole-organ-scale bioengineered grafts.
In general, current regulatory systems were primarily developed for simpler cell therapies, tissue-engineered constructs, or traditional drugs, rather than complex organ-scale bioengineered grafts[7]. The lack of standardized biosafety criteria for decellularized matrices and recellularized qualities, together with rigorous good manufacturing practice (GMP) demands, continues to hinder commercialization and delay clinical translation[7,73,81]. As a result, more practical, organ-specific regulatory frameworks are needed to better accommodate the unique biological complexities of whole-organ engineering and to accelerate its clinical application[73].
Economic burden: high development costs and limited clinical adoption
Beyond scientific and regulatory hurdles, the high financial costs associated with whole-organ bioengineering may threaten its commercial viability. The high manufacturing costs will likely restrict research and development to a few well-funded centres, slowing dissemination of the technology[63]. Further, the therapy would need to prove its worth and cost-effectiveness compared to traditional liver transplantation.
FUTURE PERSPECTIVES OF DECELLULARIZED LIVER SCAFFOLDS
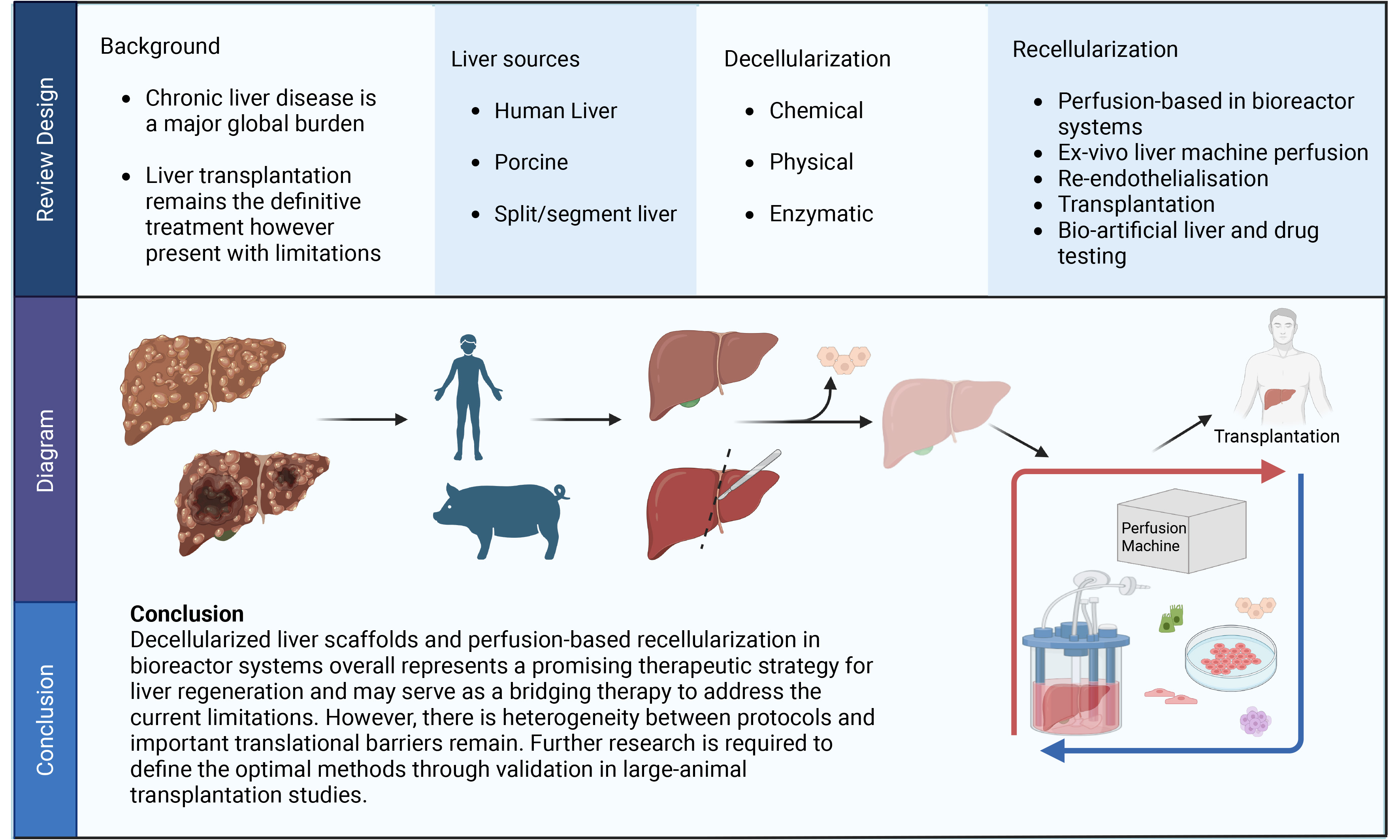
Looking ahead, decellularized liver scaffolds offer a wide range of future possibilities. Although the ultimate aim remains the development of a transplantable bioengineered liver, this goal is still far from clinical application. Emerging prospects include the adaptation of ex vivo machine perfusion principles on using partial liver segments to expand graft availability, as well as the development of bio-artificial liver and platforms for drug testing. These applications may broaden the translational and clinical relevance of liver decellularization, as illustrated in Figure 1.
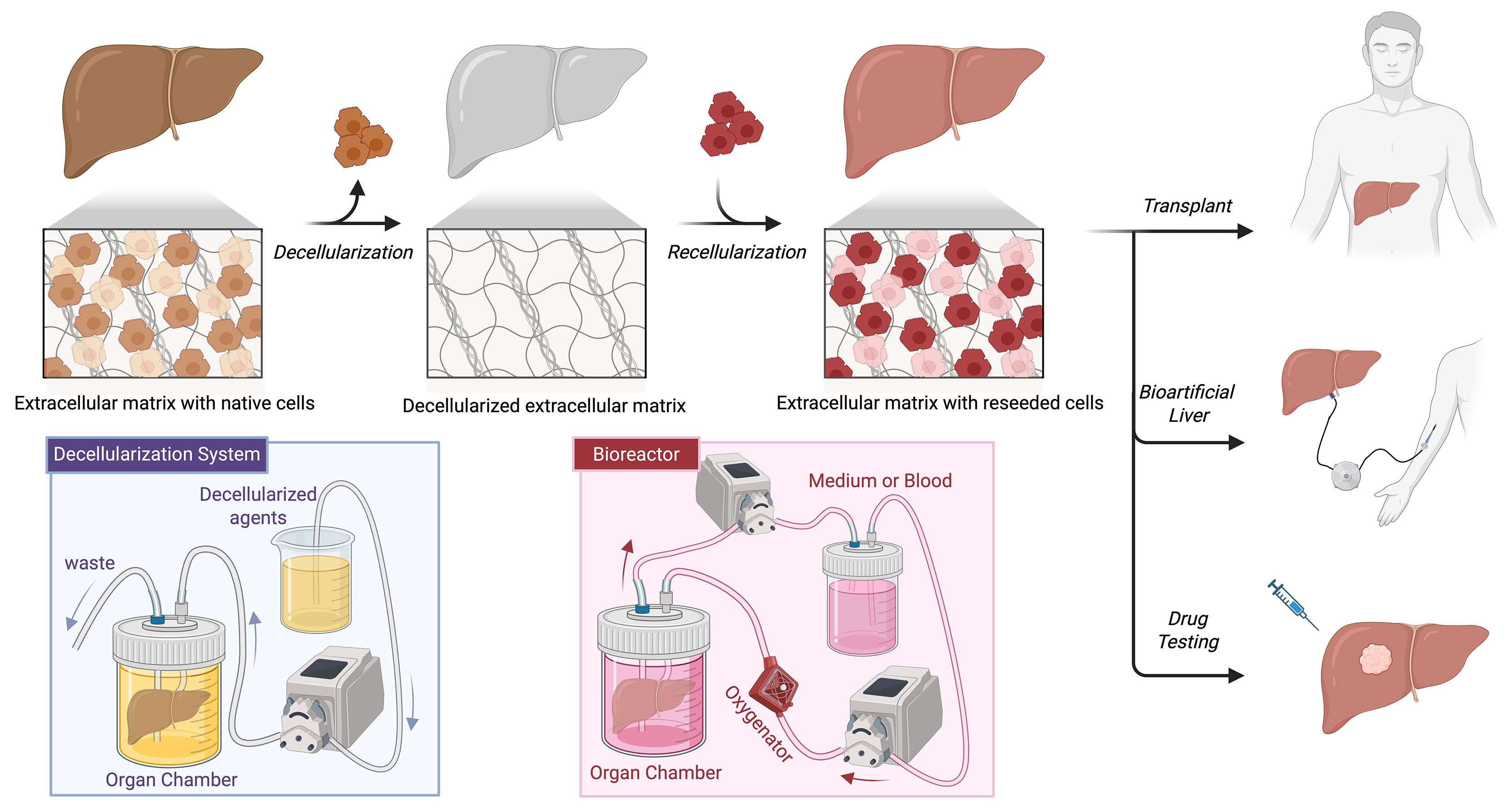
Figure 1. Schematic roadmap for decellularized liver scaffolds, perfusion-based recellularization, and future prospects. Created in BioRender. Liu, S. (2026) https://BioRender.com/kqchscx.
Split/Segmental liver scaffolds
Decellularized liver scaffolds may help address donor organ shortage by enabling modular bioengineering strategies that do not require a whole-organ graft. In the context of split/segment liver perfusion, the liver is anatomically divided into lobes and segments with independent vascular inflow and outflow, and these units may retain nearly equal metabolic functionality. Supporting this concept, Kanani et al. showed that isolated healthy liver segments from resected cancerous livers could remain viable under ex vivo machine perfusion[82]. If scaffold bioreactor perfusion can be considered an analogue of ex vivo machine perfusion, these findings point the way to a future potential. Resected liver segments from diseased or porcine liver could be decellularized and re-endothelialized, generating auxiliary grafts for partial liver support. Such an approach would increase graft availability, particularly for patients unsuitable for whole-organ transplantation.
Collectively, these lines of work suggest that coupling segmental liver scaffolds with robust vascular reconstruction may benefit patients with liver cancer or end-stage liver disease, and may also provide partial functional support for selected patients with liver cirrhosis thereby reducing metabolic demand.
Bioartificial liver and drug testing
The future scope of bioengineered liver scaffolds may also extend beyond transplantation towards bioartificial liver systems[83] and disease-modelling platforms. Liu et al. developed a perfused ex vivo human liver model using liver-specific biomatrix scaffolds and reported that the recellularized liver constructs reproduced essential hepatic functions, including metabolic and enzymatic activity[52]. Furthermore, the model was capable of simulating diclofenac-induced liver injury and generating precise hepatic responses. These findings highlight the capacity of such models in supporting predictive drug discovery, thereby broadening the scope of liver scaffold engineering beyond transplantation to preclinical drug testing.
In parallel, Chen et al. developed a tumor organoid-like tissue model using liver-derived decellularized ECM recellularized with HepG2 cells, reporting its value as a predictive platform for evaluating antitumor agents and investigating epithelial-mesenchymal transition (EMT) dynamics in tumor cells[84]. In the context of hepatocellular carcinoma, such models may provide a more physiologically relevant system for studying tumor-matrix interactions, antitumor drug toxicity, and therapeutic response in tumor progression.
CONCLUSION
Decellularized liver scaffolds represent a compelling regenerative strategy for treating cirrhosis and advanced liver disease. Although decellularization and recellularization protocols continue to improve, obstacles such as thrombogenicity after loss of the endothelial lining and the limited availability of human liver scaffolds are still being explored. Collectively, this review also highlights the need for greater emphasis on larger animal scaffolds and subsequent transplantation studies, while also pointing to the possible broader future potential of the field.
DECLARATIONS
Acknowledgments
The Graphical Abstract was created with BioRender.com [Created in BioRender. Liu, S. (2026) https://BioRender.com/qoqsawq].
Authors’ contributions
Writing - original draft, review and editing: Liu Y
Conceptualization, supervision, writing - review and editing: Liu SY, Chung WY, Bowrey D, Huang KW
Availability of data and materials
Not applicable.
AI and AI-assisted tool statement
Not applicable.
Financial support and sponsorship
None.
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Devarbhavi H, Asrani SK, Arab JP, Nartey YA, Pose E, Kamath PS. Global burden of liver disease: 2023 update. J Hepatol. 2023;79:516-37.
2. Mak LY, Liu K, Chirapongsathorn S, et al. Liver diseases and hepatocellular carcinoma in the Asia-Pacific region: burden, trends, challenges and future directions. Nat Rev Gastroenterol Hepatol. 2024;21:834-51.
3. Terrault NA, Francoz C, Berenguer M, Charlton M, Heimbach J. Liver transplantation 2023: status report, current and future challenges. Clin Gastroenterol Hepatol. 2023;21:2150-66.
4. Dai Q, Jiang W, Huang F, Song F, Zhang J, Zhao H. Recent advances in liver engineering with decellularized scaffold. Front Bioeng Biotechnol. 2022;10:831477.
5. Shaheen MF, Joo DJ, Ross JJ, et al. Sustained perfusion of revascularized bioengineered livers heterotopically transplanted into immunosuppressed pigs. Nat Biomed Eng. 2020;4:437-45.
6. Li K, Tharwat M, Larson EL, et al. Re-endothelialization of decellularized liver scaffolds: a step for bioengineered liver transplantation. Front Bioeng Biotechnol. 2022;10:833163.
7. Hussein KH, Ahmadzada B, Correa JC, et al. Liver tissue engineering using decellularized scaffolds: current progress, challenges, and opportunities. Bioact Mater. 2024;40:280-305.
8. Goldaracena N, Cullen JM, Kim D, Ekser B, Halazun KJ. Expanding the donor pool for liver transplantation with marginal donors. Int J Surg. 2020;82:30-5.
9. Plunkett J, Thinzar, King J, et al. Addressing global variation and systemic inequities in access to liver transplantation. npj Gut Liver. 2025;2:30.
10. Manzia TM, Angelico R, Gazia C, et al. De novo malignancies after liver transplantation: the effect of immunosuppression-personal data and review of literature. World J Gastroenterol. 2019;25:5356-75.
11. Magro B, Tacelli M, Mazzola A, Conti F, Celsa C. Biliary complications after liver transplantation: current perspectives and future strategies. Hepatobiliary Surg Nutr. 2021;10:76-92.
12. Mehtani R, Saigal S. Long term complications of immunosuppression post liver transplant. J Clin Hepatol. 2023;13:1103-15.
13. Yu Z, Keskinocak P, Magliocca JF, Romero R, Sokol J. Split or whole liver transplantation? Utilization and posttransplant survival. Hepatol Commun. 2023;7:e0225.
14. Lau NS, Jacques A, McCaughan G, Crawford M, Liu K, Pulitano C. Addressing the challenges of split liver transplantation through technical advances. A systematic review. Transplant Rev. 2021;35:100627.
15. Bowring MG, Massie AB, Schwarz KB, et al. Survival benefit of split‐liver transplantation for pediatric and adult candidates. Liver Transpl. 2022;28:969-82.
16. Ge J, Perito ER, Bucuvalas J, et al. Split liver transplantation is utilized infrequently and concentrated at few transplant centers in the United States. Am J Transplant. 2020;20:1116-24.
17. Groen PC, van Leeuwen OB, de Jonge J, Porte RJ. Viability assessment of the liver during ex-situ machine perfusion prior to transplantation. Curr Opin Organ Transplant. 2024;29:239-47.
18. Liew B, Nasralla D, Iype S, Pollok JM, Davidson B, Raptis DA. Liver transplant outcomes after ex vivo machine perfusion: a meta-analysis. Br J Surg. 2021;108:1409-16.
19. Op den Dries S, Karimian N, Westerkamp AC, et al. Normothermic machine perfusion reduces bile duct injury and improves biliary epithelial function in rat donor livers. Liver Transpl. 2016;22:994-1005.
20. Boehnert MU, Yeung JC, Bazerbachi F, et al. Normothermic acellular ex vivo liver perfusion reduces liver and bile duct injury of pig livers retrieved after cardiac death. Am J Transplant. 2013;13:1441-9.
21. Schliephake F, Lurje I, Uluk D, et al. Effects of hypothermic oxygenated machine perfusion on bile composition after liver transplantation - Findings from a randomized controlled trial. JHEP Rep. 2026;8:101647.
22. Ekser B, Li P, Cooper DKC. Xenotransplantation: past, present, and future. Curr Opin Organ Transplant. 2017;22:513-21.
23. Samy KP, Butler JR, Li P, Cooper DKC, Ekser B. The role of costimulation blockade in solid organ and islet xenotransplantation. J Immunol Res. 2017;2017:8415205.
24. Cross-Najafi AA, Lopez K, Isidan A, et al. Current barriers to clinical liver xenotransplantation. Front Immunol. 2022;13:827535.
25. Tao KS, Yang ZX, Zhang X, et al. Gene-modified pig-to-human liver xenotransplantation. Nature. 2025;641:1029-36.
27. Zhang W, Xu Q, Xu K, et al. Genetically engineered pig-to-human liver xenotransplantation. J Hepatol. 2026;84:587-98.
28. Mazza G, Rombouts K, Rennie Hall A, et al. Decellularized human liver as a natural 3D-scaffold for liver bioengineering and transplantation. Sci Rep. 2015;5:13079.
29. Toprakhisar B, Verfaillie CM, Kumar M. Advances in recellularization of decellularized liver grafts with different liver (stem) cells: towards clinical applications. Cells. 2023;12:301.
30. Verstegen MMA, Willemse J, van den Hoek S, et al. Decellularization of Whole Human Liver Grafts Using Controlled Perfusion for Transplantable Organ Bioscaffolds. Stem Cells Dev. 2017;26:1304-15.
31. Gao Y, Gadd VL, Heim M, et al. Combining human liver ECM with topographically featured electrospun scaffolds for engineering hepatic microenvironment. Sci Rep. 2024;14:23192.
32. Acun A, Oganesyan R, Uygun K, Yeh H, Yarmush ML, Uygun BE. Liver donor age affects hepatocyte function through age-dependent changes in decellularized liver matrix. Biomaterials. 2021;270:120689.
33. Golebiowska AA, Intravaia JT, Sathe VM, Kumbar SG, Nukavarapu SP. Decellularized extracellular matrix biomaterials for regenerative therapies: advances, challenges and clinical prospects. Bioact Mater. 2024;32:98-123.
34. Willemse J, Verstegen MMA, Vermeulen A, et al. Fast, robust and effective decellularization of whole human livers using mild detergents and pressure controlled perfusion. Mater Sci Eng C Mater Biol Appl. 2020;108:110200.
35. Jiang Y, Li R, Han C, Huang L. Extracellular matrix grafts: from preparation to application (Review). Int J Mol Med. 2021;47:463-74.
36. Kasravi M, Ahmadi A, Babajani A, et al. Immunogenicity of decellularized extracellular matrix scaffolds: a bottleneck in tissue engineering and regenerative medicine. Biomater Res. 2023;27:10.
37. Moffat D, Ye K, Jin S. Decellularization for the retention of tissue niches. J Tissue Eng. 2022;13:20417314221101151.
38. Kim S, Park SH, Mun J, et al. Enhancing peripheral nerve regeneration throughNaOH‐based decellularization of human nerve tissue. Bioeng Transl Med. 2025;10:e70072.
39. Neishabouri A, Soltani Khaboushan A, Daghigh F, Kajbafzadeh AM, Majidi Zolbin M. Decellularization in tissue engineering and regenerative medicine: evaluation, modification, and application methods. Front Bioeng Biotechnol. 2022;10:805299.
40. Koo MA, Jeong H, Hong SH, Seon GM, Lee MH, Park JC. Preconditioning process for dermal tissue decellularization using electroporation with sonication. Regen Biomater. 2021;9:rbab071.
41. Ansari T, Southgate A, Obiri-Yeboa I, et al. Development and characterization of a porcine liver scaffold. Stem Cells Dev. 2020;29:314-26.
42. Kasturi M, Vasanthan KS. Harvesting decellularized liver extracellular matrix from rodents for 3D scaffold fabrication. Artif Cells Nanomed Biotechnol. 2024;52:175-85.
43. Weigl EJ, Demir S, Schmid T, Hotes A, Muensterer O, Kappler R. Establishing a three-dimensional scaffold model of hepatoblastoma. Front Bioeng Biotechnol. 2023;11:1229490.
44. Rabbani M, Zakian N, Alimoradi N. Contribution of physical methods in decellularization of animal tissues. J Med Signals Sens. 2021;11:1-11.
45. Zhang Y, Lv Y, Wang Y, Chang TT, Rubinsky B. Pancreatic islets implanted in an irreversible electroporation generated extracellular matrix in the liver. Radiol Oncol. 2023;57:51-8.
46. Stefan L, Uehara MK, Ikegami Y, et al. Early-stage transplant responses in miniature-liver implants based on decellularization-recellularization technology. J Biosci Bioeng. 2025;140:244-51.
47. Ijima H, Fukuda Y, Uehara MK, et al. Decellularized mouse liver as a small-scale scaffold for the creation of a miniaturized human liver. J Chem Eng Japan. 2023;56:2204899.
48. Alaby Pinheiro Faccioli L, Suhett Dias G, Hoff V, et al. Optimizing the decellularized porcine liver scaffold protocol. Cells Tissues Organs. 2022;211:385-94.
49. Anderson BD, Nelson ED, Joo D, et al. Functional characterization of a bioengineered liver after heterotopic implantation in pigs. Commun Biol. 2021;4:1157.
50. Higashi H, Yagi H, Kuroda K, et al. Transplantation of bioengineered liver capable of extended function in a preclinical liver failure model. Am J Transplant. 2022;22:731-44.
51. Horie H, Oshima Y, Fukumitsu K, et al. Antithrombotic revascularization strategy of bioengineered liver using a biomimetic polymer. Tissue Eng Part A. 2025;31:433-41.
52. Liu J, Hanson A, Yin W, et al. Decellularized liver scaffolds for constructing drug-metabolically functional ex vivo human liver models. Bioact Mater. 2024;43:162-80.
53. Afzal Z, Huguet EL. Bioengineering liver tissue by repopulation of decellularised scaffolds. World J Hepatol. 2023;15:151-79.
54. Afrin S, Yadav U, Yadav CJ, Kamel J, Lee JY, Park KM. Development of an enhanced liver scaffold recellularization using fibronectin. J Biomater Appl. 2025;40:513-28.
55. Panahi F, Baheiraei N, Sistani MN, Salehnia M. Analysis of decellularized mouse liver fragment and its recellularization with human endometrial mesenchymal cells as a candidate for clinical usage. Prog Biomater. 2022;11:409-20.
56. Kang SH, Kim MY, Eom YW, Baik SK. Mesenchymal stem cells for the treatment of liver disease: present and perspectives. Gut Liver. 2020;14:306-15.
57. Takeishi K, Collin de l’Hortet A, Wang Y, et al. Assembly and function of a bioengineered human liver for transplantation generated solely from induced pluripotent stem cells. Cell Rep. 2020;31:107711.
58. Acun A, Oganesyan R, Jaramillo M, Yarmush ML, Uygun BE. Human-origin iPSC-based recellularization of decellularized whole rat livers. Bioengineering. 2022;9:219.
59. Kern S, Eichler H, Stoeve J, Klüter H, Bieback K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells. 2009;24:1294-301.
60. Kadota Y, Yagi H, Inomata K, et al. Mesenchymal stem cells support hepatocyte function in engineered liver grafts. Organogenesis. 2014;10:268-77.
61. Chen J, Ma S, Yang H, et al. Generation and metabolomic characterization of functional ductal organoids with biliary tree networks in decellularized liver scaffolds. Bioact Mater. 2023;26:452-64.
62. Sassi L, Ajayi O, Campinoti S, et al. A perfusion bioreactor for longitudinal monitoring of bioengineered liver constructs. Nanomaterials. 2021;11:275.
63. Nelson VL, Stumbras AR, Palumbo RN, et al. Manufacturing and Functional Characterization of Bioengineered Liver Grafts for Extracorporeal Liver Assistance in Acute Liver Failure. Bioengineering. 2023;10:1201.
64. Campinoti S, Almeida B, Goudarzi N, et al. Rat liver extracellular matrix and perfusion bioreactor culture promote human amnion epithelial cell differentiation towards hepatocyte-like cells. J Tissue Eng. 2023;14:20417314231219813.
65. Yadav CJ, Yadav U, Afrin S, Lee JY, Kamel J, Park KM. Heparin immobilization enhances hemocompatibility, re-endothelization, and angiogenesis of decellularized liver scaffolds. Int J Mol Sci. 2024;25:12132.
66. Guo B, Zhou Q, Chen J, et al. Orthotopic transplantation of functional bioengineered livers in rats. ACS Biomater Sci Eng. 2023;9:1940-51.
67. Dias ML, Paranhos BA, Goldenberg RCDS. Liver scaffolds obtained by decellularization: a transplant perspective in liver bioengineering. J Tissue Eng. 2022;13:20417314221105305.
68. Zahmatkesh E, Khoshdel Rad N, Hossein-Khannazer N, et al. Cell and cell-derivative-based therapy for liver diseases: current approaches and future promises. Expert Review of Gastroenterol Hepatol. 2023;17:237-49.
69. Chen W, Lv L, Chen N, Cui E. Immunogenicity of mesenchymal stromal/stem cells. Scand J Immunol. 2023;97:e13267.
70. Zhao T, Zhang ZN, Rong Z, Xu Y. Immunogenicity of induced pluripotent stem cells. Nature. 2011;474:212-5.
71. Otsuka R, Wada H, Murata T, Seino KI. Immune reaction and regulation in transplantation based on pluripotent stem cell technology. Inflamm Regen. 2020;40:12.
72. Yoon J, Lee S, Kim MJ, Kim JH. Brief summary of the regulatory frameworks of regenerative medicine therapies. Front Pharmacol. 2025;15:1486812.
73. van Hengel EVA, van der Laan LJW, de Jonge J, Verstegen MMA. Towards safety and regulation criteria for clinical applications of decellularized organ-derived matrices. Bioengineering. 2025;12:136.
74. Framework for the regulation of regenerative medicine products. Available from: https://www.fda.gov/vaccines-blood-biologics/cellular-gene-therapy-products/framework-regulation-regenerative-medicine-products. [Last accessed on 9 Jun 2026].
75. Fang WH, Vangsness CT Jr. Governmental regulations and increasing food and drug administration oversight of regenerative medicine products: what’s new in 2020? Arthroscopy. 2020;36:2765-70.
76. Detela G, Lodge A. EU regulatory pathways for ATMPs: standard, accelerated and adaptive pathways to marketing authorisation. Mol Ther Methods Clin Dev. 2019;13:205-32.
77. European Medicines Agency. Advanced therapy medicinal products: overview. Available from: https://www.ema.europa.eu/en/human-regulatory-overview/advanced-therapy-medicinal-products-overview. [Last accessed on 9 Jun 2026].
78. Pharmaceuticals and Medical Devices Agency. Medical devices. Available from: https://www.pmda.go.jp/english/pnavi_e-06.html. [Last accessed on 9 Jun 2026].
79. Maria Cristina Galli. Regulatory aspects of gene therapy and cell therapy products: a global perspective. 2nd ed. Springer Cham; 2023. Available from: https://doi.org/10.1007/978-3-031-34567-8. [Last accessed on 9 Jun 2026].
80. Yokomizo R, Hosoya S, Umezawa A, Okamoto A. Amendment of the Act on the Safety of Regenerative Medicine: a new voyage of regenerative medicine in Japan and the dawn of a new era for young investigators. Regen Ther. 2025;29:237-46.
81. Silva DN, Chrobok M, Ahlén G, Blomberg P, Sällberg M, Pasetto A. ATMP development and pre-GMP environment in academia: a safety net for early cell and gene therapy development and manufacturing. Immunooncol Technol. 2022;16:100099.
82. Kanani T, Isherwood J, Chung WY, et al. A O03 ex vivo perfusion of isolated human liver segments: the development of a novel model for ethical, translational research. Br J Surg. 2022;109:znac404.003.
83. Wu G, Wu D, Lo J, et al. A bioartificial liver support system integrated with a DLM/GelMA-based bioengineered whole liver for prevention of hepatic encephalopathy via enhanced ammonia reduction. Biomater Sci. 2020;8:2814-24.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].