


Genetic variation in non-coding RNAs contributes to the biological mechanism of cardiovascular diseases
 0
0 Abstract
Cardiovascular diseases (CVDs) involve structural and functional abnormalities of the heart and blood vessels, in which genetic factor plays a significant role. Non-coding RNAs (ncRNAs), as important products of genetic material, are important contributors to CVDs. They mainly regulate the expression of targets and participate in various biological processes such as cell proliferation, cell apoptosis, and signal transduction. Genetic variation in ncRNAs can affect the expression levels of ncRNAs themselves and their downstream targets, leading to the dysregulation of biological processes and contributing to the occurrence and development of CVDs. This review aims to summarize the current research status of ncRNA genetic variation in CVDs and clarify the functions and biological mechanisms of ncRNA genetic variation involved in CVDs.
Keywords
INTRODUCTION
The cardiovascular diseases (CVDs), a leading cause of morbidity and mortality worldwide, encompass a range of disorders affecting the heart and blood vessels, such as ischemic cardiomyopathy, non-ischemic cardiomyopathy, and heart failure[1]. CVDs are characterized primarily by structural and functional alterations in the heart or blood vessels. The pathogenesis of CVDs is influenced by multiple factors, with genetic factor playing a dominant role. Non-coding RNAs (ncRNAs), which constitute nearly 98% of genetic material, do not encode protein but are key regulators of gene expression[2]. The impact of ncRNA variation on CVDs has attracted increasing attention.
Based on length, ncRNAs can be classified into microRNA (miRNA), PIWI-interacting RNA (piRNA), circular RNA (circRNA), long non-coding RNA (lncRNA), and others. ncRNAs regulate gene expression by targeting mRNAs to inhibit protein synthesis, participate in epigenetic regulation of chromatin, and also engage in mutual regulatory networks among themselves. For example, both lncRNAs and circRNAs can act as miRNA sponges, which capture miRNAs and prevent their binding to target mRNAs[3-5].
The type of ncRNA genetic variation includes single nucleotide polymorphisms (SNPs) and indels. SNP, a point genetic variation caused by base substitution, represents the most common variation in ncRNA. Genetic variation in ncRNAs may alter their own biogenesis, including the changes in expression levels and the formation of different isoforms[6,7]. Moreover, ncRNAs genetic variation can affect the binding to their targets, because of the alterations in secondary structures and nucleotide changes in target binding regions, even the creation of new binding sites[8,9]. Consequently, ncRNA genetic variation can disrupt normal regulatory functions, thereby promoting or suppressing the progression of CVDs.
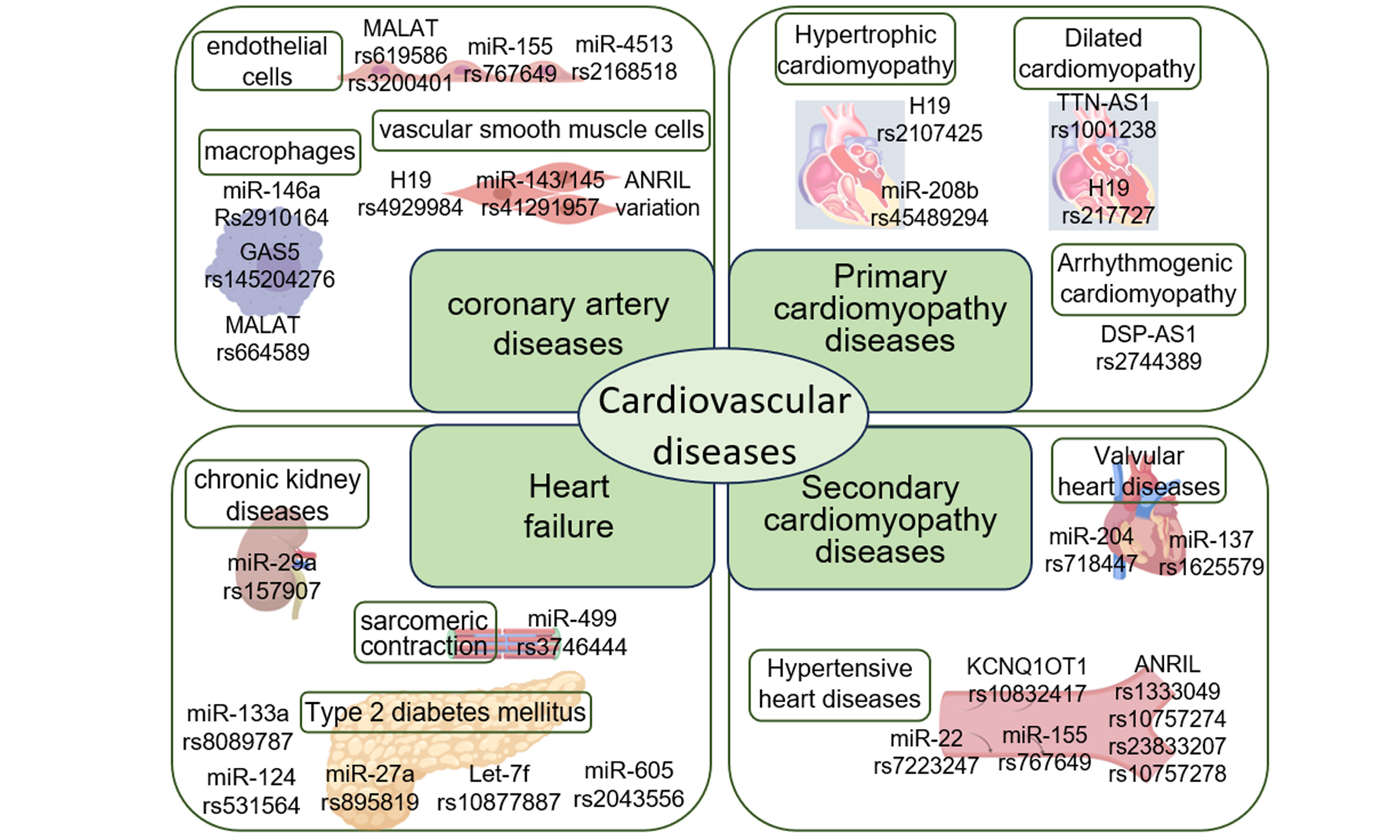
The associations between ncRNA variation and CVDs have been reported in numerous studies. Herein, we summarize currently documented ncRNA variations implicated in CVDs [Table 1], and elaborate on selected examples. This review focuses on the critical roles of ncRNA genetic variation in the pathogenesis of CVDs, overviews and discusses the pathogenic effects and mechanisms through which ncRNA genetic variation contributes to CVDs. A literature search has been performed on PubMed, Web of Science by selecting one keyword from each of the following three groups and combining them for search. Group 1 includes “non-coding RNA”, “micro-RNA”, “lncRNA”, “circRNA”, “piRNA”, group 2 consists of disease names including “cardiovascular diseases”, “coronary artery diseases”, “hypertrophic cardiomyopathy”, “dilated cardiomyopathy” and so on, and group 3 includes “variation”, “mutation”, “SNP”, etc. There is no date restriction in writing review article.
ncRNA variations in CVDs
| Name | ncRNA type | Disease | SNP variation | Reference |
| ANRIL | lncRNA | Coronary artery disease | rs10757278, rs7341786, rs7341791 | [10] |
| rs10811650, rs496892, rs564398, rs6475606 | [11] | |||
| rs1333049 | [12] | |||
| Hypertensive heart disease | rs10757274, rs2383206, rs10757278, rs1333049 | [13] | ||
| DSP-AS1 | lncRNA | Arrhythmogenic cardiomyopathy | rs2744389 | [14] |
| FENDRR/FOXF1 | lncRNA | Hypertrophic cardiomyopathy | rs40384, rs39529, rs39527 | [15] |
| GAS5 | lncRNA | Coronary artery disease | rs145204276 | [16] |
| H19 | lncRNA | Coronary artery disease | rs4929984 | [17] |
| Hypertrophic cardiomyopathy | rs2107425 | [18] | ||
| Dilated cardiomyopathy | rs217727 | [19] | ||
| KCNQ1OT1 | lncRNA | Hypertensive heart disease | rs10832417, rs3782064, | [8] |
| let-7f | lncRNA | Heart failure | rs10877887 | [20] |
| Linc-VWF | lncRNA | Coronary artery disease | rs1558324 | [21] |
| LINC00305 | lncRNA | Coronary artery disease | rs2850711 | [22] |
| rs2676671 | [23] | |||
| MALAT1 | lncRNA | Coronary artery disease | rs619586 | [24] |
| rs664589 | [25] | |||
| mir1-2 | miRNA | Hypertrophic cardiomyopathy | rs9989532 | [26] |
| miR-133 | miRNA | Hypertrophic cardiomyopathy | 79T>A | [27] |
| miR-133a | miRNA | Heart failure | rs8089787 | [20] |
| miR-137 | miRNA | Valvular heart disease | rs1625579 | [28] |
| miR-143/miR-145 | miRNA | Coronary artery disease | rs41291957 | [6] |
| mir-146a | miRNA | Coronary artery disease | rs2910164, rs2431697 | [29] |
| miR-155 | miRNA | Hypertensive heart disease | rs767649 | [30] |
| miR-196a | miRNA | Coronary artery disease | rs11614913 | [31] |
| miR-22 | miRNA | Hypertensive heart disease | rs7223247 | [32] |
| miR-29a | miRNA | Heart failure | rs157907 | [33] |
| miR-204 | miRNA | Valvular heart disease | rs718447 | [34] |
| miR-208b | miRNA | Hypertrophic cardiomyopathy | rs45489294 | [26] |
| mir-367 | miRNA | Hypertrophic cardiomyopathy | rs13136737 | [26] |
| mir-499 | miRNA | Heart failure | u17c | [35] |
| rs3746444 | [36] | |||
| mir-590 | miRNA | Hypertrophic cardiomyopathy | rs6971711 | [37] |
| TTN-AS1 | lncRNA | Dilated cardiomyopathy | rs1001238 | [38] |
NCRNA GENETIC VARIATION AND ISCHEMIC CARDIOMYOPATHY
Ischemic cardiomyopathy mainly refers to coronary artery diseases (CADs), which is caused by myocardial ischemia resulting from coronary atherosclerosis. The pathological process involves multiple cell types, such as endothelial cells, smooth muscle cells, and macrophages[39]. ncRNAs coordinate complex molecular networks in CADs and regulate various biological processes including lipid metabolism, inflammatory response, cell proliferation, and cell apoptosis[40]. Instead, ncRNA variations disrupt their regulatory function.
Dysregulated lipid metabolism and the consequent increase in plasma lipoprotein levels represent a major etiological factor in CADs, particularly involving abnormalities of low-density lipoprotein (LDL)[41]. Numerous studies have shown association between ncRNA variations and dyslipidemia. For example, miR-155 inhibits the cyclic adenosine monophosphate/protein kinase A (cAMP/PKA) signaling pathway then promoting lipid metabolism[42], and has been reported as a novel biomarker of CAD[43]. Rs767649 can influence its expression as a common variation of miR-155. A research based on a Chinese Tibetan population shows that carriers of the miR-155 rs767649 variant exhibit significantly higher serum miR-155 levels and lipid concentrations compared to controls[30]. Cholesterol as a major lipid plays a crucial role in the pathogenesis of CADs. The lncRNA MALAT1 can promote cholesterol accumulation via regulating the miR-17-5p/ABCA1 axis[44]. MALAT1 rs619586 A>G variant reported in a Chinese population is significantly associated with a decrease in total cholesterol, and may play a protective role[24]. The MALAT1 rs3200401 variant in a Chinese Han population has similar function to influence the total cholesterol[45]. Furthermore, ncRNAs can influence lipid levels by targeting and regulating genes related to lipid metabolism. But variation disrupting this regulation may lead to CADs. For instance, the miR-4513 rs2168518 G>A variant reduces the expression of miR-4513, resulting in increased levels of its target genes MTMR3 and BNC2 both of which are associated with lipid metabolism, thereby affecting serum LDL and total cholesterol levels[46].
On the other hand, elevated blood lipid levels promote the deposition of LDL in blood vessels, triggering vascular inflammation[47]. The inflammatory response induces endothelial dysfunction and macrophage infiltration, further accelerating the progression of CADs. The activation of inflammation is strictly regulated, while ncRNA variation may disrupt this regulation. The miR-146a has been proved as an anti-inflammatory factor in many research[48,49]. But the miR-146a rs2910164 G>C variant leads to decrease of its expression and overactivation of inflammation in atrial fibrillation that results in the occurrence of ischemic events[50]. And this variant prevents miR-146a from binding to the 3’UTR of the IKBA gene, leading to activation of the NF-κB inflammatory pathway and accelerating CADs[51]. Another variant rs2431697 of miR-146a also was reported to be associated with a higher risk of CADs in a Chinese population by influencing its expression and function[29]. The lncRNA LINC00305 regulates anti-inflammatory factor miR-124 as a miRNA sponge[52]. A research shows that individuals carrying the LINC00305 rs2850711 A>T variant have higher activation levels of NF-κB and inflammation than controls[22]. We also find a CAD-related variant rs2676671 of LINC00305 reported in ncRNAVar database[23], but the underlying mechanisms remain uninvestigated. Furthermore, variation can disrupt these interactions of ncRNAs engaging in mutual regulatory networks, thereby influencing inflammatory responses. For instance, the lncRNA MALAT1 rs664589 C>G variant alters the regulatory capacity of MALAT1 toward miRNAs, including miR-1972, miR-194, miR-4717, and others then increasing the risk of ischemic events in a Chinese population[25]. In CADs, as the barrier of vessel, endothelial cells not only exhibit dysfunction caused by inflammation, also show dysregulation of its proliferation and apoptosis. ncRNA variations may play a significant role in this dysregulation. The lncRNA GAS5 rs145204276 variant affects endothelial proliferation and apoptosis via GAS5/miR-21/PDCD4 pathway, associated with the risk of CADs[16].
The proliferation and phenotypic switching of vascular smooth muscle cells (VSMCs) also play crucial roles in the progression of CADs. The lncRNA ANRIL, an antisense ncRNA located at the locus of the cyclin-dependent kinase inhibitor (CDKN2a/b) gene, inhibits the expression of CDKN2a/b and then regulates cell proliferation. The ANRIL rs1333049 variant is an independent determinant factor of CADs in Chinese population[12]. Meanwhile, a study on stroke in Chinese population found that this variant increases ANRIL expression, which enhances the inhibitory effect on CDKN2a/b and promotes the proliferation of VSMCs[53]. Therefore, the ANRIL rs1333049 variant likely contributes to the progression of CADs by affecting the proliferation of VSMCs. Conversely, ANRIL G>A variants in rs7341786 or rs7341791 result in increased formation of circular ANRIL (circANRIL) which loses its inhibitory effect on CDKN2a/b, reduces cell proliferation, and exerts a protective role in CADs[10]. Besides, ncRNAs regulate cell proliferation-related signaling pathways, and variations in these ncRNAs can lead to abnormal proliferation. For example, the lncRNA H19 rs4929984 C>A variant upregulates H19 expression level, which promotes VSMC proliferation and inhibits apoptosis via the MAPK and NF-κB signaling pathways[54]. Also, phenotypic switching of VSMCs is a key process in CADs. The miR-143/miR-145 targets a network of transcription factors to promote VSMC differentiation and proliferation[55]. But the miR-143/miR-145 rs41291957 G>A variant, located at upstream of miR-143, upregulates miR-143/miR-145 expression, inhibits the phenotypic switching of VSMCs, and thus exhibits a protective effect in CADs[6].
NCRNA GENETIC VARIATION AND NON-ISCHEMIC CARDIOMYOPATHY
Non-ischemic cardiomyopathy, unlike ischemic cardiomyopathy, is characterized by the absence of ischemic injury. According to the main affected organs or systems, it is categorized into primary cardiomyopathy and secondary cardiomyopathy. Primary cardiomyopathy is confined to or predominantly involves the myocardium, whereas secondary cardiomyopathy manifests as pathological cardiomyopathy changes that form part of numerous systemic disorders and typically involve multiple organs[56]. ncRNAs also play regulatory roles in non-ischemic cardiomyopathy, and alterations in their regulatory functions caused by ncRNA genetic variation can influence the progression of non-ischemic cardiomyopathy.
ncRNA genetic variation and primary cardiomyopathy
Primary cardiomyopathy can be further classified into hypertrophic cardiomyopathy, dilated cardiomyopathy, arrhythmogenic cardiomyopathy, left ventricular noncompaction cardiomyopathy, among others. This section focuses on the role of ncRNA variations in hypertrophic cardiomyopathy, dilated cardiomyopathy, and arrhythmogenic cardiomyopathy.
ncRNA genetic variation and hypertrophic cardiomyopathy
Hypertrophic cardiomyopathy (HCM) is the most common inherited structural heart disease, characterized by left ventricular hypertrophy. The core pathogenesis is sarcomeric dysfunction , which manifests as hypercontractility, leading to cardiomyocyte hypertrophy, disarray, and myocardial fibrosis. Approximately 60% of HCM patients carry pathogenic variations in sarcomeric protein genes, such as beta-myosin heavy chain (MYH7)[57] and myosin binding protein (MYBPC3)[58]. NcRNAs directly regulate the expression of sarcomeric proteins, variations in these ncRNAs may disrupt such regulation to play a significant role in HCM progression. For example, miR-208b directly binds to SOX-6, a transcriptional repressor of MYH7, thereby upregulating the expression of MYH7[59]. Meanwhile, study in the Czech population has shown that the frequency of miR-208b rs45489294 C>A variant is significantly higher in HCM patients compared to healthy individuals[26]. This suggests that miR-208b variation may predispose carriers to HCM by influencing MYH7 expression, although direct experimental confirmation is still lacking.
Calcium ions are key regulators of muscle contraction and relaxation, making calcium handling particularly crucial in cardiomyocytes. ncRNAs are involved in regulating calcium handling proteins, while variation can alter this regulatory function, thereby inhibiting or promoting calcium signal. For example, the lncRNA H19 acts as a sponge for miR-675, inhibiting miR-675 targeting to the ion channel phosphorylation protein CaMKIID, thus influencing ion channel regulation and calcium handling[60]. In a study about HCM patients from northern Spain, the H19 rs2107425 T>C variant was found to be more prevalent in the patient group with higher H19 expression levels[18]. This suggests that H19 variation may affect calcium handling by regulating the phosphorylation status of ion channels, though this remains to be experimentally validated. Additionally, ncRNAs can directly act on proteins involved in calcium signaling, but ncRNA variations may lead to dysregulation. For instance, calcineurin, a key mediator of calcium signaling, interacts with miR-133 to mutually regulate their expression levels[61]. The 79T>A point variation in miR-133 gene has been identified on atrial fibrillation, which alters the processing of miR-133, leading to the accumulation of the normally degraded miR-133-5p and consequent changes in regulated targets[27]. This suggests that miR-133 variant in HCM may induce calcineurin levels and disrupt calcium signaling.
Overload of cardiomyocyte sarcomeric proteins can lead to hypertrophy and fibrosis which is the key processes of HCM. ncRNAs are involved in regulating cardiac hypertrophy and fibrosis, and variation can disturb these regulations. For example, miR-590 can regulate the transforming growth factor β (TGF-β), Activin and protein kinase B (AKT) signaling pathways, thereby participating in the modulation of cardiac phenotypes. The miR-590 rs6971711 C>T variant in the African-American population, results in decreased levels of mature miR-590 without affecting the expression of pri-miR-590 and pre-miR-590, and decreases the inhibitory capacity of miR-590 on its targets, promoting cardiomyocyte hypertrophy and fibrosis, thus contributing to HCM[37].
ncRNA genetic variation and dilated cardiomyopathy
Dilated cardiomyopathy (DCM) is characterized by left ventricular dilation and persistent systolic dysfunction in the absence of abnormal loading conditions or CADs. In contrast to HCM, which is characterized by increased contractility but impaired relaxation, DCM is primarily defined by impaired myocardial contractility. Similar to HCM, sarcomeric protein genes are major pathogenic genes in DCM, such as MYH7[62], titin (TTN)[63]. Besides, DCM-associated pathogenic genes also include cytoskeletal protein genes, mitochondrial homeostasis-related genes, such as filamin C (FLNC)[64] and dnaJ heat shock protein family member C19 (DNAJC19)[65]. Correspondingly, ncRNA variation disrupts the direct regulatory effects of ncRNAs on sarcomeric proteins. For example, lncRNA TTN-AS1 is an antisense transcript of TTN gene, which can bind to TTN mRNA and regulate TTN expression[66]. In addition, the TTN-AS1 rs1001238 T>C variant on skeletal muscle was found to alter its own structure, which may exacerbate muscle damage and inflammation by affecting molecules related to sarcomeric structure[67]. This suggests that TTN-AS1 variation may promote the development of DCM by altering the regulation of TTN, leading to changes in sarcomeric structure.
Mitochondrial function can be impaired by mutations in mitochondrial homeostasis-related genes and sarcomeric or cytoskeletal protein genes, leading to insufficient energy supply for cardiomyocytes and exacerbating DCM progression. ncRNAs are involved in the regulation of mitochondrial function, and variation in ncRNAs can result in dysregulation. For example, the lncRNA cytochrome B (CYTB), located at cardiomyocyte mitochondria, can act as a sponge for miR-103 to regulate reactive oxygen species (ROS) generation, thereby participating in the modulation of mitochondrial function[68]. However, currently no reports indicate the effects of ncRNA variation on mitochondrial function in DCM. Besides, the lncRNA H19 inhibits anti-apoptosis gene PA2G4 via targeting miR-675 to promote cardiomyocyte apoptosis, which accelerates the progression of DCM[69]. The rs217727 variant of H19 reported in a German population is significantly associated with DCM, because variant may disrupt its secondary structure and lead to key region failing to bind targets[19]. This suggests that the H19 variation changes its structure and the interaction with its targets, then playing an important role in DCM via influencing the regulation of apoptosis.
ncRNA genetic variation and arrhythmogenic cardiomyopathy
Arrhythmogenic cardiomyopathy (ACM) is characterized that myocardium is replaced by the fibrofatty infiltration, along with cardiomyocyte loss and inflammation. ACM is an inherited cardiomyopathy that most of the mutations causing ACM is located at genes encoding desmosomal proteins, such as desmoplakin (DSP)[70], so ACM is also named desmosomal disease. Compared with ncRNA, its variation may loss the function to regulate desmosomal proteins. For instance, lncRNA DSP-AS1 rs2744389 variant reported in an Italy population participates in ACM via a DSP-independent pathway, because DSP-AS1 can regulate DSP mRNA and protein levels but the variant not[14].
Furthermore, the instability of intercellular junctions caused by desmosomal proteins is associated with the Wnt and Hippo signaling pathways. ncRNAs can participate in ACM by targeting these pathways, but the variation disrupts. For instance, the lincRNA-p21 targets β-catenin involving in the Wnt signaling pathway, then playing a regulatory role in ACM[71,72]. Additionally, lncRNA RP11-323N12.5 and circYap regulate the transcriptional initiation and mRNA stability of YAP1, respectively, thereby participating in the Hippo pathway[73,74]. However, no related variations have been reported to date.
ncRNA genetic variation and secondary cardiomyopathy
Secondary cardiomyopathy is typically systemic disorders. This section focuses on the role of ncRNA variations in hypertensive heart diseases and valvular heart diseases.
ncRNA genetic variation and hypertensive heart diseases
Hypertensive heart diseases (HHDs) are characterized by cardiac structural or functional impairment caused by long-term hypertension, involving left ventricular hypertrophy, systolic and diastolic dysfunction, and other pathological changes. Numerous ncRNAs have been implicated in the pathogenesis of HHDs. Left ventricular hypertrophy in HHDs is primarily attributed to cardiomyocyte proliferation and fibrosis. miR-1[75], lncRNA ERBB4-IR[76] and circ_0005372[77] have all been reported to participate in biological processes such as cardiomyocyte proliferation, migration, and myocardial fibrosis in HHDs.
The miR-22 rs7223247 variant, located at downstream of miR-22, plays a significant role in HHDs via affecting left ventricular mass reported in a British Caucasian population[32]. And miR-22 has been proven to be associated with HCM in animal models, by regulating the expression levels of calcium ion transporter protein SERCA2A and muscle-restricted genes to influence calcium homeostasis and myofibrillar protein content[78]. This suggests that miR-22 variation may disrupt calcium homeostasis and decrease myofibrillar protein content then aggravating HHDs. Besides, there are various ncRNA variations in HHDs focusing on their association with disease susceptibility. For example, the lncRNA KCNQ1OT1 has been reported to promote HHDs via targeting miR-301b and Tcf7[79]. And the KCNQ1OT1 rs10832417 variant may alter its secondary structure and its binding to target miRNAs, resulting in decrease of KCNQ1OT1 expression, potentially reducing the risk of hypertension[8]. The lncRNA GAS5 participates in HHD development by regulating miR‑21, activating the PI3K/AKT signaling pathway and its downstream targets[80]. Meanwhile, the GAS5 rs145204276 variant located in the promoter region changes the binding site of transcriptional factor and resulting in higher levels of GAS5[81]. This suggests that the variation in GAS5 may alter its expression levels and downstream signaling pathway, thereby contributing to HHDs, although there are no evidences to date. However, significant gaps remain in our understanding of ncRNA variations in HHDs. For instance, study has shown that lncRNA ANRIL variants (rs10757274, rs2383207, rs10757278, and rs1333049) are all associated with the risk of HHDs[13]. Similarly, the miR-155 rs767649 variant was reported to be associated with susceptibility of HHDs[30]. But, the precise mechanisms by which these ncRNA variation contributes to HHDs remain unclear and require further investigation.
ncRNA genetic variation and valvular heart diseases
Valvular heart diseases (VHDs) primarily affect the mitral and aortic valves, and to a lesser extent the tricuspid and pulmonary valves. VHDs are characterized by extensive tissue remodeling, including calcification, endothelial dysfunction[82]. Calcific aortic valve diseases (CAVDs) are the most prevalent among VHDs. Cardiac valves consist of valvular endothelial cells (VECs) and valvular interstitial cells (VICs), and the differentiation of VICs into osteoblast-like cells is a hallmark of valvular calcification. ncRNAs in VHDs are involved in regulating biological processes such as tissue remodeling, like circRNA circARHGAP10 and miR-25 both of which target osteogenic differentiation and influence CAVDs[83,84]. MiR-204 has been identified as a core regulatory factor in CAVDs and occupies a central position in the regulatory network. Its expression is modulated by various lncRNAs such as TUG1 and MALAT1, and it regulates osteogenic differentiation by directly targeting the TGF‑β/BMP2 pathway and downstream genes like RUNX2 and SMAD4[85,86]. Study has shown that the rs718447 A>G variant in the upstream flanking region of miR-204, decreases its expression[34]. This suggests that miR-204 variant may influence its expression, disrupt its regulatory functions and influence VHD progression, although there are no reports linking miR-204 variant to VHDs yet. Additionally, miR-137 participates in endothelial osteogenic differentiation, as a downstream target of lncRNA OIP5-AS1[87]. MiR-137 rs1625579 variant is located at seed sequence and downregulates miR-137 expression levels[28]. This suggested that miR-137 variation may change its expression levels to influence endothelial osteogenic differentiation, and then play a role in VHD.
NCRNA GENETIC VARIATION AND HEART FAILURE
Heart failure (HF) is characterized by impaired cardiac blood-pumping capacity caused by structural and/or functional abnormalities of the heart, which fails to pump enough blood to meet tissue metabolic demands. HF involves diverse etiologies, risk factors, and comorbidities[88,89]. Given that most CVDs manifests as HF at the terminal stage, most ncRNAs associated with HF have already been elucidated in other CVDs. For instance, miR-499 regulates sarcomeric contraction via Myh7b[90]and has been investigated in ACM[91], HCM[92], and VHDs[93]. Furthermore, plasma levels of miR‑499 are significantly elevated in HF patients[94]. This finding suggested that miR-499 may participate in HF pathogenesis. Consistent with this, the U17C variant located at the 3’ end of miR-499(outside the seed sequence), induces miR-499 to aberrantly recruit target genes into the RNA-induced silencing complex (RISC), thereby improving cardiac function via modulating the homeostasis of cardiac mRNAs and proteins[35]. Similarly, miR- 499 rs3746444 A>G variant affects its maturation process, changing anti-apoptosis function and influencing HF[36].
On the other hand, HF represents the end stage of not only CVDs but also various metabolic diseases, including type 2 diabetes mellitus (T2DM) and chronic kidney diseases (CKDs). NcRNA variations play a significant role in this kind of HF. MiR-133a regulates Akt signaling pathway, to improve cardiac function and inhibit HF[95]. But a study about an Italy population demonstrated that miR-133a rs8089787 variant leads to cardiac diastolic dysfunction, which enhance the progression of HF in patients with T2DM[20]. MiRNA let-7a targeting TGFBR3 regulates p38 MAPK signaling pathway and apoptosis, playing a significant role in HF[96]. However, the let-7f rs10877887 variant was reported to increase the risk of HF in T2DM patients[20]. MiR-27a was reported to inhibit mitophagy mediated by PINK1/Parkin pathway via targeting AMPKα2[20], and miR-124 targeting CD151 negatively regulates cardiac angiogenesis and cardiac function in HF[97]. But, the research in an Italy population shows that the miR-27a rs895819 variant as a protective effect, and the miR-124 rs531564 as a risk effect, are associated with HF via influencing the progression of T2DM[88]. The miR-146a can regulate MMP2/16-MLCK3-p-MLC2 contractile axis increasing the risk of HF and the miR-146a rs2910164 variant is associated with T2DM, which may influence HF reported in a Pakistani population[98-100]. The miR-605 regulates TDRG1/TNFRSF21 influencing TGF-β signaling pathway and the miR-605 rs2043556 variant also was reported to affect the regulation of mature miRNAs and mRNA targets, then playing a significant role in T2DM, which may regulate HF[101,102]. Otherwise, CKDs are another metabolic disorder causing HF. The miR-29a was reported to target PPARδ signaling pathway, and the miR-29a rs157907 A>G variant may compromise its own stability, thereby being significantly associated with a high CKD risk, which means a high HF risk[33,103].
DISCUSSION
The pathogenesis of CVDs exhibits a dual nature of complexity: it arises from the involvement of multiple biological processes within a single disease entity and is compounded by the sharing of common pathological mechanisms among distinct cardiovascular disorders. In the same disease, variants in a ncRNA can participate in different biological processes, like the lncRNA MALAT1. In CADs, the MALAT1 rs619586 variant influences total cholesterol, while MALAT1 rs664589 variant affects inflammatory response[24,25]. This phenomenon may be caused by ncRNA variants with different secondary structures and targets, also variants may have different functions in different type of cells. Moreover, the same biological process is involved in distinct CVDs. Sarcomeric proteins are essential for heart muscle and their abnormalities can lead to CVDs, like HCM[104], DCM[105], HF[106]. But the difference lies in the distinct sarcomeric proteins involved in different diseases. MiR-208b variation directly targets the sarcomeric protein MYH7 in HCM[26], TTN-AS1 variation targets the sarcomeric protein TTN in DCM[67], and miR-499 variation regulates sarcomeric protein Myh7b in HF[36]. Calcium, as key signaling molecules in muscle cells, their concentration in the cytoplasm is strictly regulated. Abnormalities of calcium can affect cardiomyocyte contraction, leading to HCM[107], DCM[108], HHD[109], HF[110]. H19 variation regulates calcium/calmodulin-dependent protein kinase II (CaMKIID) to influence calcium concentration, leading to HCM[18]. In HHD, miR-22 variation can regulate calcium transport protein SERCA2A[32].
The widespread distribution of ncRNAs across different cell types means that the same ncRNA may be involved in distinct cardiovascular diseases. For example, the lncRNA ANRIL rs1333049 variant promotes disease progression in both CADs[111] and HHDs[13]. Similarly, the lncRNA H19 rs217727 variant increases the risk of both CADs[112] and DCM[19]. This may be due to the fact that they participate in the same biological processes across different diseases and perform analogous functions, but further research is still needed to reveal the underlying mechanisms. On the other hand, the same ncRNA variation may exert completely opposite effects in different populations. For instance, when miR-146a rs2910164 is substituted to G, it is associated with increased CAD risk in the Turkish population[113], whereas it confers reduced risk in the Chinese population[114]. Although this opposite effect is attributed to population-specific, the underlying mechanisms remain to be fully elucidated. Extending beyond our focus on specific cardiovascular diseases, evidence suggests that ncRNA variation participates in a broader spectrum of cardiovascular disorders. For example, the miR-26 rs2910164 and rs4636297 variants are the risk factors for acute myocardial infarction in the Pakistani population[115]. Similarly, the miR-618 rs2682818 variant increases the risk of ischemic stroke[116].
NcRNA variation influences disease pathogenesis mainly through two aspects: effects on their own expression and alterations in their regulatory functions. The effects on expression are manifested as follows: (a) Variation in transcriptional regulatory regions such as enhancers, silencers, or promoters (e.g. the lncRNA PCAT19 rs11672691 variant is located at an enhancer element region and contributes to prostate cancer by altering the binding site of the transcription factor HOXA2[117]); (b) copy number variations (e.g. an increased copy number of lncRNA LINC01133 is associated with poor prognosis in hepatocellular carcinoma[118]); (c) Effect on the maturation process, especially miRNA (e.g. miR-27a rs11671784 variant disrupts the maturation process from pre-miR-27a to mature miR-27a, decreasing the risk of gastric cancer[119]; (d) altered isoform formation (e.g. the lncRNA ANRIL rs10757278 variant results in the generation of isoform circANRIL[10]); (e) structural changes (e.g. the lncRNA KCNQ1OT1 rs10832417 variant alters secondary structure and is correlated with HHDs[8]. Alteration in regulatory functions: (a) Changes in binding to target miRNAs, especially miRNA sponge lncRNA and circRNA (e.g. the base G in lncRNA TINCR rs2288947 is the target binding of miR-1247, while the base C in TINCR rs8113645 becomes the binding site of miR-30c[120]); (b) Changes in binding to target mRNAs (e.g. miR-146a rs2910164 is located at the binding site, which alters the binding to IKBA[9,51]); (c) Alterations in binding to target proteins [e.g. lncRNA antihypertrophic interrelated transcript (AHIT)[121] and lncRNA UC.323[122] both directly bind proteins involved in histone methylation, suggesting that variation in AHIT and UC.323 could disrupt epigenetic regulation]. Besides, by studying how ncRNA variations affect themselves, we can elucidate the effects of primary structure changes on secondary structure, knowledge which in turn informs the study of coding RNA mutations. Simultaneously, investigation of the ncRNA variations provides insights into the disease mechanisms involving the ncRNAs, highlighting its critical role.
Research on ncRNA variations can discover the pathogenesis of diseases. The lncRNA ANRIL, identified from a CVD risk locus on chromosome 9p21.3, can promote atherosclerosis via miR-181b-5p-TMEM106B-NHE1-lysosomal pH axis[123-125]. Many variations in ANRIL have been proven to be associated with CVDs, like rs1333049, rs7341786, rs7341791, which perfects the elucidation of disease mechanisms[10,12]. Besides, study on ncRNA variations provides a new approach for identifying biomarkers associated with CVDs. The lncRNA IGBP1P1 was identified as a key regulator of cardiomyocyte size and cardiac function via association analysis between variations and potential target genes[126]. However, most research has focused on ncRNAs, and research on variation remains limited. With the development of next-generation sequencing technologies and structural prediction models, studies on the functional variations of ncRNAs will continue to increase in the future. On the other hand, ncRNA variations are the potential candidates of targeted therapy. Currently, most of targeted therapies based on ncRNA rely on siRNA or ASO-mediated mechanisms, both of which exert therapeutic effects by silencing target genes. For example, Inclisiran(ALN-PCSsc) is a siRNA targeting proprotein convertase subtilisin-kexin type 9 (PCSK9), to treat atherosclerosis by reducing LDL-cholesterol levels[127]. Studies on ncRNA variations indicate that therapeutic efficacy can be attained by generating siRNA or ASO variations with altered target gene specificity, which may serve as a promising research direction and therapeutic strategy in the future. Moreover, TP53 Y220C variation has been used to treat cancer, via restoring TP53 suppressor function by a compound rezatapopt[128]. This suggests that specific compounds targeting ncRNA variants can be identified and screened for therapeutic applications according to ncRNA variations.
CONCLUSION
This review summarizes current research on ncRNA variation in CVDs and their impacts on related biological mechanisms. However, much of the existing work focuses on associations between ncRNA variants and disease risk, often without clarifying the functional consequences or precise pathogenic mechanisms. Investigating how ncRNA variants alter ncRNA biogenesis, target gene regulation, and signaling pathway activation will enhance our understanding of cardiovascular disease pathogenesis and may facilitate the discovery of novel diagnostic biomarkers and therapeutic targets.
DECLARATIONS
Acknowledgment
The Graphical Abstract was created with BioGDP.com.
Authors’ contributions
Conceptualization, writing, and supervision: Tu X
Data curation and writing: Wang X
Both authors contributed to the article and approved the submitted version.
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tool Doubao (version 1.8, released 2025-12-18) was used solely for language editing. The tool did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
None.
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Martin SS, Aday AW, Almarzooq ZI, et al. ; American Heart Association Council on Epidemiology and Prevention Statistics Committee and Stroke Statistics Subcommittee. 2024 heart disease and stroke statistics: a report of US and Global Data From the American Heart Association. Circulation. 2024;149:e347-913.
2. International Human Genome Sequencing Consortium. Finishing the euchromatic sequence of the human genome. Nature. 2004;431:931-45.
3. O'Brien J, Hayder H, Zayed Y, Peng C. Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol. 2018;9:402.
4. Zhang X, Wang W, Zhu W, et al. Mechanisms and functions of long non-coding RNAs at multiple regulatory levels. Int J Mol Sci. 2019;20:5573.
5. Samra M, Srivastava K. Non-coding RNA and their potential role in cardiovascular diseases. Gene. 2023;851:147011.
6. Hall IF, Climent M, Viviani Anselmi C, et al. rs41291957 controls miR-143 and miR-145 expression and impacts coronary artery disease risk. EMBO Mol Med. 2021;13:e14060.
7. Holdt LM, Teupser D. Long noncoding RNA ANRIL: lnc-ing genetic variation at the chromosome 9p21 locus to molecular mechanisms of atherosclerosis. Front Cardiovasc Med. 2018;5:145.
8. Xie Y, Qi H, Peng W, et al. SNPs in lncRNA KCNQ1OT1 modulate its expression and confer susceptibility to salt sensitivity of blood pressure in a Chinese han population. Nutrients. 2022;14:3990.
9. Mihajlovic M, Savic-veselinovic M, Farkic M, Zeljic K. MiR-146a gene variant rs2910164 might be associated with coronary in-stent restenosis risk: results from a pilot study and meta-analysis. Genetika. 2022;54:933-46.
10. Burd CE, Jeck WR, Liu Y, Sanoff HK, Wang Z, Sharpless NE. Expression of linear and novel circular forms of an INK4/ARF-associated non-coding RNA correlates with atherosclerosis risk. PLoS Genet. 2010;6:e1001233.
11. Congrains A, Kamide K, Oguro R, et al. Genetic variants at the 9p21 locus contribute to atherosclerosis through modulation of ANRIL and CDKN2A/B. Atherosclerosis. 2012;220:449-55.
12. Li Q, Peng W, Li H, Zhuang J, Luo X, Xu Y. Association of the single nucleotide polymorphism in chromosome 9p21 and chromosome 9q33 with coronary artery disease in Chinese population. BMC Cardiovasc Disord. 2017;17:255.
13. Bayoglu B, Yuksel H, Cakmak HA, Dirican A, Cengiz M. Polymorphisms in the long non-coding RNA CDKN2B-AS1 may contribute to higher systolic blood pressure levels in hypertensive patients. Clin Biochem. 2016;49:821-7.
14. Foco L, De Bortoli M, Del Greco M F, et al. Genomic and molecular evidence that the LncRNA DSP-AS1 modulates desmoplakin expression. Hum Genet. 2025;144:843-60.
15. Cuesta-Llavona E, Lorca R, Rolle V, et al. Association of the genetic variation in the long non-coding RNA FENDRR with the risk of developing hypertrophic cardiomyopathy. Life. 2022;12:818.
16. Shen Z, She Q. Association between the deletion allele of Ins/Del polymorphism (Rs145204276) in the promoter region of GAS5 with the risk of atherosclerosis. Cell Physiol Biochem. 2018;49:1431-43.
17. Huang J, Li M, Li J, et al. LncRNA H19 rs4929984 variant is associated with coronary artery disease susceptibility in han chinese female population. Biochem Genet. 2021;59:1359-80.
18. Gómez J, Lorca R, Reguero JR, et al. Genetic variation at the long noncoding RNA H19 gene is associated with the risk of hypertrophic cardiomyopathy. Epigenomics. 2018;10:865-73.
19. Martens L, Rühle F, Witten A, et al. A genetic variant alters the secondary structure of the lncRNA H19 and is associated with dilated cardiomyopathy. RNA Biol. 2021;18:409-15.
20. Sun D, Zhu Z, Zhang Y, et al. Relation of genetic polymorphisms in microRNAs with diastolic and systolic function in type 2 diabetes mellitus. Nutr Metab Cardiovasc Dis. 2022;32:2877-82.
21. Liu Y, Ferguson JF, Xue C, et al. Tissue-specific RNA-Seq in human evoked inflammation identifies blood and adipose LincRNA signatures of cardiometabolic diseases. Arterioscler Thromb Vasc Biol. 2014;34:902-12.
22. Wahba AS, Ibrahim ME, Mesbah NM, Saleh SM, Abo-Elmatty DM, Mehanna ET. Serum LINC00305 expression and its genetic variant rs2850711 are associated with clinical and laboratory features of rheumatoid arthritis. Br J Biomed Sci. 2020;77:142-7.
23. Zhang W, Zeng B, Yang M, et al. ncRNAVar: a manually curated database for identification of noncoding RNA variants associated with human diseases. J Mol Biol. 2021;433:166727.
24. Wang G, Li Y, Peng Y, Tang J, Li H. Association of polymorphisms in MALAT1 with risk of coronary atherosclerotic heart disease in a Chinese population. Lipids Health Dis. 2018;17:75.
25. Yu H, Tu S, Shen C, et al. Effects of the lncRNA MALAT1 gene region rs664589 site mutation on acute myocardial infarction in Chinese han. Cell Mol Biol. 2024;70:119-27.
26. Curila K, Benesova L, Tomasov P, et al. Variants in miRNA regulating cardiac growth are not a common cause of hypertrophic cardiomyopathy. Cardiology. 2015;130:137-42.
27. Ohanian M, Humphreys DT, Anderson E, Preiss T, Fatkin D. A heterozygous variant in the human cardiac miR-133 gene, MIR133A2, alters miRNA duplex processing and strand abundance. BMC Genet. 2013;14:18.
28. Mahmoud RH, Hefzy EM, Shaker OG, et al. GAS5 rs2067079 and miR-137 rs1625579 functional SNPs and risk of chronic hepatitis B virus infection among Egyptian patients. Sci Rep. 2021;11:20014.
29. Wang Y, Wang X, Li Z, et al. Two single nucleotide polymorphisms (rs2431697 and rs2910164) of miR-146a are associated with risk of coronary artery disease. Int J Environ Res Public Health. 2017;14:514.
30. Yan W, Chen R, Zhao Y, Zhang X, Li X. MicroRNA-155-5p rs767649 polymorphism is associated with susceptibility to essential hypertension in the Chinese Tibetan population in the Gannan Area. Artery Res. 2025;31:70.
31. Fragoso JM, Ramírez-Bello J, Martínez-Ríos MA, et al. miR-196a2 (rs11614913) polymorphism is associated with coronary artery disease, but not with in-stent coronary restenosis. Inflamm Res. 2019;68:215-21.
32. Harper AR, Mayosi BM, Rodriguez A, et al. Common variation neighbouring micro-RNA 22 is associated with increased left ventricular mass. PLoS One. 2013;8:e55061.
33. Mirani Sargazi F, Alidadi A, Taheri H, et al. Functional miR29a gene polymorphism enhanced the risk of chronic kidney disease in an Iranian population: a preliminary case-control study and bioinformatics analyses. Meta Gene. 2020;25:100755.
34. Butrym A, Łacina P, Kuliczkowski K, Bogunia-Kubik K, Mazur G. Genetic variation of the gene coding for microRNA-204 (miR-204) is a risk factor in acute myeloid leukaemia. BMC Cancer. 2018;18:107.
35. Dorn GW 2nd, Matkovich SJ, Eschenbacher WH, Zhang Y. A human 3’ miR-499 mutation alters cardiac mRNA targeting and function. Circ Res. 2012;110:958-67.
36. Ding W, Li M, Sun T, et al. A polymorphism rs3746444 within the pre-miR-499 alters the maturation of miR-499-5p and its antiapoptotic function. J Cell Mol Med. 2018;22:5418-28.
37. Lin X, Steinberg S, Kandasamy SK, et al. Common miR-590 Variant rs6971711 Present only in African Americans reduces miR-590 biogenesis. PLoS One. 2016;11:e0156065.
38. Kucher AN, Nazarenko MS. Regulatory potential of noncoding RNAs Co-located with cardiomyopathy related genes. Russ J Genet. 2023;59:325-43.
39. Zaman S, Wasfy JH, Kapil V, et al. The Lancet Commission on rethinking coronary artery disease: moving from ischaemia to atheroma. Lancet. 2025;405:1264-312.
40. Abbas M, Gaye A. Emerging roles of noncoding RNAs in cardiovascular pathophysiology. Am J Physiol Heart Circ Physiol. 2025;328:H603-21.
41. Borén J, Chapman MJ, Krauss RM, et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease: pathophysiological, genetic, and therapeutic insights: a consensus statement from the European Atherosclerosis Society Consensus Panel. Eur Heart J. 2020;41:2313-30.
42. Gu X, Liu H, Luo W, Wang X, Wang H, Li L. Di-2-ethylhexyl phthalate-induced miR-155-5p promoted lipid metabolism via inhibiting cAMP/PKA signaling pathway in human trophoblastic HTR-8/Svneo cells. Reprod Toxicol. 2022;114:22-31.
43. Qiu XK, Ma J. Alteration in microRNA-155 level correspond to severity of coronary heart disease. Scand J Clin Lab Invest. 2018;78:219-23.
44. Liu L, Tan L, Yao J, Yang L. Long non‑coding RNA MALAT1 regulates cholesterol accumulation in ox‑LDL‑induced macrophages via the microRNA‑17‑5p/ABCA1 axis. Mol Med Rep. 2020;21:1761-70.
45. Li Y, Zhang D, Zhang Y, et al. Association of lncRNA polymorphisms with triglyceride and total cholesterol levels among myocardial infarction patients in Chinese population. Gene. 2020;724:143684.
46. Ghanbari M, de Vries PS, de Looper H, et al. A genetic variant in the seed region of miR-4513 shows pleiotropic effects on lipid and glucose homeostasis, blood pressure, and coronary artery disease. Hum Mutat. 2014;35:1524-31.
47. Duewell P, Kono H, Rayner KJ, et al. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature. 2010;464:1357-61.
48. Zhang L, Wang IC, Meng S, Xu J. miR-146a Decreases Inflammation and ROS Production in Aged Dermal Fibroblasts. Int J Mol Sci. 2024;25:6821.
49. Yang X, Ren Y, Li X, Xia L, Wan J. MiR-146a reduces inflammation in experimental pancreatitis via the TRAF6-NF-κB signaling pathway in mice. Immun Inflamm Dis. 2025;13:e70163.
50. Roldán V, Arroyo AB, Salloum-Asfar S, et al. Prognostic role of MIR146A polymorphisms for cardiovascular events in atrial fibrillation. Thromb Haemost. 2014;112:781-8.
51. Qiao XR, Zheng T, Xie Y, et al. MiR-146a rs2910164 (G/C) polymorphism is associated with the development and prognosis of acute coronary syndromes: an observational study including case control and validation cohort. J Transl Med. 2023;21:325.
52. Li X, Yu M, Han L, et al. LINC00305 represses miR-124 expression to trigger inflammatory insults in the presence of lipopolysaccharide. Artif Cells Nanomed Biotechnol. 2019;47:2352-60.
53. Yang J, Gu L, Guo X, et al. LncRNA ANRIL expression and ANRIL gene polymorphisms contribute to the risk of ischemic stroke in the Chinese han population. Cell Mol Neurobiol. 2018;38:1253-69.
54. Pan JX. LncRNA H19 promotes atherosclerosis by regulating MAPK and NF-kB signaling pathway. European Review for Medical and Pharmacological Sciences. 2017;21:322-8. Available from: https://www.europeanreview.org/wp/wp-content/uploads/322-328-LncRNA-H19-promotes-atherosclerosis-by-promoting-the-expression-of-p38-and-p65.pdf [Last accessed on 1 Jun 2026].
55. Rangrez AY, Massy ZA, Metzinger-Le Meuth V, Metzinger L. miR-143 and miR-145: molecular keys to switch the phenotype of vascular smooth muscle cells. Circ Cardiovasc Genet. 2011;4:197-205.
56. Wang Y, Jia H, Song J. Accurate classification of non-ischemic cardiomyopathy. Curr Cardiol Rep. 2023;25:1299-317.
57. Lee S, Vander Roest AS, Blair CA, et al. Incomplete-penetrant hypertrophic cardiomyopathy MYH7 G256E mutation causes hypercontractility and elevated mitochondrial respiration. Proc Natl Acad Sci U S A. 2024;121:e2318413121.
58. Greer-Short A, Greenwood A, Leon EC, et al. AAV9-mediated MYBPC3 gene therapy with optimized expression cassette enhances cardiac function and survival in MYBPC3 cardiomyopathy models. Nat Commun. 2025;16:2196.
59. Kim JM, Lim KS, Hong JS, Kang JH, Lee YS, Hong KC. A polymorphism in the porcine miR-208b is associated with microRNA biogenesis and expressions of SOX-6 and MYH7 with effects on muscle fibre characteristics and meat quality. Anim Genet. 2015;46:73-7.
60. Liu L, An X, Li Z, et al. The H19 long noncoding RNA is a novel negative regulator of cardiomyocyte hypertrophy. Cardiovasc Res. 2016;111:56-65.
61. Dong DL, Chen C, Huo R, et al. Reciprocal repression between microRNA-133 and calcineurin regulates cardiac hypertrophy: a novel mechanism for progressive cardiac hypertrophy. Hypertension. 2010;55:946-52.
62. Naderi N, Mohsen-Pour N, Nilipour Y, Pourirahim M, Maleki M, Kalayinia S. A novel heterozygous missense MYH7 mutation potentially causes an autosomal dominant form of myosin storage myopathy with dilated cardiomyopathy. BMC Cardiovasc Disord. 2023;23:487.
63. Ghahremani S, Kanwal A, Pettinato A, et al. CRISPR activation reverses haploinsufficiency and functional deficits caused by TTN truncation variants. Circulation. 2024;149:1285-97.
64. Verdonschot JAJ, Vanhoutte EK, Claes GRF, et al. A mutation update for the FLNC gene in myopathies and cardiomyopathies. Hum Mutat. 2020;41:1091-111.
65. Janz A, Chen R, Regensburger M, et al. Generation of two patient-derived iPSC lines from siblings (LIBUCi001-A and LIBUCi002-A) and a genetically modified iPSC line (JMUi001-A-1) to mimic dilated cardiomyopathy with ataxia (DCMA) caused by a homozygous DNAJC19 mutation. Stem Cell Res. 2020;46:101856.
66. Zheng QX, Wang J, Gu XY, et al. TTN-AS1 as a potential diagnostic and prognostic biomarker for multiple cancers. Biomed Pharmacother. 2021;135:111169.
67. Pimenta EM, Santos CGMD, Assis MGD, Veneroso CE, Soalheiro I, Serpa TKF. TTN-AS1 genotype (rs1001238) and its influence on inflammatory responses in muscle tissues in soccer players. Braz arch biol technol. 2024;67:e24231067.
68. Zhang X, Yuan S, Liu J, et al. Overexpression of cytosolic long noncoding RNA cytb protects against pressure-overload-induced heart failure via sponging microRNA-103-3p. Mol Ther Nucleic Acids. 2022;27:1127-45.
69. Zhang Y, Zhang M, Xu W, Chen J, Zhou X. The long non-coding RNA H19 promotes cardiomyocyte apoptosis in dilated cardiomyopathy. Oncotarget. 2017;8:28588-94.
70. Heliö K, Brandt E, Vaara S, et al. DSP c.6310delA p.(Thr2104Glnfs*12) associates with arrhythmogenic cardiomyopathy, increased trabeculation, curly hair, and palmoplantar keratoderma. Front Cardiovasc Med. 2023;10:1130903.
71. Yoon JH, Abdelmohsen K, Srikantan S, et al. LincRNA-p21 suppresses target mRNA translation. Mol Cell. 2012;47:648-55.
72. Piquer-Gil M, Domenech-Dauder S, Sepúlveda-Gómez M, Machí-Camacho C, Braza-Boïls A, Zorio E. Non coding RNAs as regulators of Wnt/β-catenin and hippo pathways in arrhythmogenic cardiomyopathy. Biomedicines. 2022;10:2619.
73. Wang J, Huang F, Shi Y, et al. RP11-323N12.5 promotes the malignancy and immunosuppression of human gastric cancer by increasing YAP1 transcription. Gastric Cancer. 2021;24:85-102.
74. Wu N, Yuan Z, Du KY, et al. Translation of yes-associated protein (YAP) was antagonized by its circular RNA via suppressing the assembly of the translation initiation machinery. Cell Death Differ. 2019;26:2758-73.
75. Connolly M, Garfield BE, Crosby A, Morrell NW, Wort SJ, Kemp PR. miR-1-5p targets TGF-βR1 and is suppressed in the hypertrophying hearts of rats with pulmonary arterial hypertension. PLoS One. 2020;15:e0229409.
76. Li JC, Jia J, Dong L, et al. Angiotensin II mediates hypertensive cardiac fibrosis via an Erbb4-IR-dependent mechanism. Mol Ther Nucleic Acids. 2023;33:180-90.
77. Zhu M, Li G, Li Y, Chen J. Circ_0005372 targets the miR-153-3p/ITGB3 axis to stimulate the PI3K/AKT signaling pathway to facilitate the occurrence and development of congenital heart disease and pulmonary arterial hypertension in children. Cardiovasc Pathol. 2026;80:107768.
78. Gurha P, Abreu-Goodger C, Wang T, et al. Targeted deletion of microRNA-22 promotes stress-induced cardiac dilation and contractile dysfunction. Circulation. 2012;125:2751-61.
79. E M, Ren F, Yu Y, Li H, Shen C. The role of lncRNAKCNQ1OT1/miR-301b/Tcf7 axis in cardiac hypertrophy. Cardiol Young. 2024;34:1493-505.
80. Zheng D, Hou Y, Li Y, et al. Long non-coding RNA Gas5 is associated with preeclampsia and regulates biological behaviors of trophoblast via MicroRNA-21. Front Genet. 2020;11:188.
81. Tang Y, Wang Y, Wang X, Liu Y, Zheng K. A genetic variant of rs145204276 in the promoter region of long noncoding RNA GAS5 is associated with a reduced risk of breast cancer. Clin Breast Cancer. 2019;19:e415-21.
82. Lampropoulos K, Penteris M, Gerotziafas G. Direct oral anticoagulants in valvular diseases and prosthetic valves: why not? Cardiovasc Drugs Ther. 2026;40:697-709.
83. Jiang Y, Zhu J, Chen Z, et al. CircRNA ARHGAP10 promotes osteogenic differentiation through the miR-335-3p/RUNX2 pathway in aortic valve calcification. J Thorac Dis. 2023;15:5971-91.
84. Yu C, Wu D, Zhao C, Wu C. CircRNA TGFBR2/MiR-25-3p/TWIST1 axis regulates osteoblast differentiation of human aortic valve interstitial cells. J Bone Miner Metab. 2021;39:360-71.
85. Yu C, Li L, Xie F, et al. LncRNA TUG1 sponges miR-204-5p to promote osteoblast differentiation through upregulating Runx2 in aortic valve calcification. Cardiovasc Res. 2018;114:168-79.
86. Xiao X, Zhou T, Guo S, et al. LncRNA MALAT1 sponges miR-204 to promote osteoblast differentiation of human aortic valve interstitial cells through up-regulating Smad4. Int J Cardiol. 2017;243:404-12.
87. Zheng D, Wang B, Zhu X, et al. LncRNA OIP5-AS1 inhibits osteoblast differentiation of valve interstitial cells via miR-137/TWIST11 axis. Biochem Biophys Res Commun. 2019;511:826-32.
88. McDonagh TA, Metra M, Adamo M, et al. ; ESC Scientific Document Group. 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure: Developed by the Task Force for the diagnosis and treatment of acute and chronic heart failure of the European Society of Cardiology (ESC) With the special contribution of the Heart Failure Association (HFA) of the ESC. Eur Heart J. 2021;42:3599-726.
89. Triposkiadis F, Xanthopoulos A, Parissis J, Butler J, Farmakis D. Pathogenesis of chronic heart failure: cardiovascular aging, risk factors, comorbidities, and disease modifiers. Heart Fail Rev. 2022;27:337-44.
90. Yeung F, Chung E, Guess MG, Bell ML, Leinwand LA. Myh7b/miR-499 gene expression is transcriptionally regulated by MRFs and Eos. Nucleic Acids Res. 2012;40:7303-18.
91. Calore M, Lorenzon A, Vitiello L, et al. A novel murine model for arrhythmogenic cardiomyopathy points to a pathogenic role of Wnt signalling and miRNA dysregulation. Cardiovasc Res. 2019;115:739-51.
92. Baulina N, Pisklova M, Kiselev I, Chumakova O, Zateyshchikov D, Favorova O. Circulating miR-499a-5p is a potential biomarker of MYH7-associated hypertrophic cardiomyopathy. Int J Mol Sci. 2022;23:3791.
93. Gupta SK, Kumari S, Singh S, Barthwal MK, Singh SK, Thum T. Non-coding RNAs: regulators of valvular calcification. J Mol Cell Cardiol. 2020;142:14-23.
94. Corsten MF, Dennert R, Jochems S, et al. Circulating MicroRNA-208b and MicroRNA-499 reflect myocardial damage in cardiovascular disease. Circ Cardiovasc Genet. 2010;3:499-506.
95. Sang HQ, Jiang ZM, Zhao QP, Xin F. MicroRNA-133a improves the cardiac function and fibrosis through inhibiting Akt in heart failure rats. Biomed Pharmacother. 2015;71:185-9.
96. Chen CY, Choong OK, Liu LW, et al. MicroRNA let-7-TGFBR3 signalling regulates cardiomyocyte apoptosis after infarction. EBioMedicine. 2019;46:236-47.
97. Zhao Y, Yan M, Chen C, et al. MiR-124 aggravates failing hearts by suppressing CD151-facilitated angiogenesis in heart. Oncotarget. 2018;9:14382-96.
98. Dong M, Chen D, Zhu Y, et al. Impaired regulation of MMP2/16-MLCK3 by miR-146a-5p increased susceptibility to myocardial ischaemic injury in aging mice. Cardiovasc Res. 2023;119:786-801.
99. Ciccacci C, Di Fusco D, Cacciotti L, et al. MicroRNA genetic variations: association with type 2 diabetes. Acta Diabetol. 2013;50:867-72.
100. Khan MS, Rahman B, Haq TU, et al. Deciphering the variants located in the MIR196A2, MIR146A, and MIR423 with type-2 diabetes mellitus in Pakistani population. Genes. 2021;12:664.
101. Cheng J, Tang Y, Cai X, Gong J. Long noncoding RNAs testis development related gene 1 aggravates transforming growth factor-β1-induced fibrogenesis and inflammatory response of cardiac fibroblasts via miR-605-3p/tumor necrosis factor receptor superfamily-21 axis. J Cardiovasc Pharmacol. 2022;79:296-303.
102. Moudi M, Montazer Zohour M. SNP in microRNA sequences or binding sites of miRNAs: association with type 2 diabetes mellitus susceptibility and in silico analysis. Int J Diabetes Dev Ctries. 2018;38:330-7.
103. Zhang S, Yin Z, Dai FF, et al. miR-29a attenuates cardiac hypertrophy through inhibition of PPARδ expression. J Cell Physiol. 2019;234:13252-62.
104. Argirò A, Parikh V, Jurcut R, et al. Hypertrophic cardiomyopathy. Nat Rev Dis Primers. 2025;11:58.
105. Schultheiss HP, Fairweather D, Caforio ALP, et al. Dilated cardiomyopathy. Nat Rev Dis Primers. 2019;5:32.
106. Hamdani N, Kooij V, van Dijk S, et al. Sarcomeric dysfunction in heart failure. Cardiovasc Res. 2008;77:649-58.
107. Viola HM, Hool LC. Impaired calcium handling and mitochondrial metabolic dysfunction as early markers of hypertrophic cardiomyopathy. Arch Biochem Biophys. 2019;665:166-74.
108. Wang Y, Zhou T, Zhao J, et al. Calcium handling remodeling in dilated cardiomyopathy: From molecular mechanisms to targeted therapies. Channels. 2025;19:2519545.
109. Wang JG, Palmer BF, Vogel Anderson K, Sever P. Amlodipine in the current management of hypertension. J Clin Hypertens. 2023;25:801-7.
110. Redel-Traub G, Marx SO, Marks AR. Targeting calcium regulation for heart failure and arrhythmia therapeutics: a critical review. Circulation. 2025;152:957-70.
111. Letonja J, Petrovič D, Petrovič D. CDKN2B-AS1 polymorphism rs1333049 is associated with advanced carotid artery atherosclerosis in a Slovenian population. Biomol Biomed. 2025;25:1362-8.
112. Gao W, Zhu M, Wang H, et al. Association of polymorphisms in long non-coding RNA H19 with coronary artery disease risk in a Chinese population. Mutat Res. 2015;772:15-22.
113. Oner T, Arslan C, Yenmis G, et al. Association of NFKB1A and microRNAs variations and the susceptibility to atherosclerosis. J Genet. 2017;96:251-9.
114. Bao Q, Li R, Wang C, et al. Association between microRNA-146a rs2910164 polymorphism and coronary heart disease: an updated meta-analysis. Medicine. 2022;101:e31860.
115. Ali S, Haq TU, Hussain M, et al. Genetic variants rs2910164, rs4636297 and rs895819 may contribute to the onset of acute myocardial infarction in Pakistani population. PLoS One. 2024;19:e0296025.
116. Zhang Z, Xu G, Cai B, Zhang H, Zhu W, Liu X. Genetic variants in MicroRNAs predict recurrence of ischemic stroke. Mol Neurobiol. 2017;54:2776-80.
117. Gao P, Xia JH, Sipeky C, et al. ; PRACTICAL Consortium. Biology and clinical implications of the 19q13 aggressive prostate cancer susceptibility locus. Cell. 2018;174:576-589.e18.
118. Yin D, Hu ZQ, Luo CB, et al. LINC01133 promotes hepatocellular carcinoma progression by sponging miR-199a-5p and activating annexin A2. Clin Transl Med. 2021;11:e409.
119. Yang Q, Jie Z, Ye S, et al. Genetic variations in miR-27a gene decrease mature miR-27a level and reduce gastric cancer susceptibility. Oncogene. 2014;33:193-202.
120. Xu C, Liu M, Jia D, Tao T, Hao D. lncRNA TINCR SNPs and expression levels are associated with bladder cancer susceptibility. Genet Test Mol Biomarkers. 2021;25:31-41.
121. Yu J, Yang Y, Xu Z, et al. Long noncoding RNA ahit protects against cardiac hypertrophy through SUZ12 (suppressor of zeste 12 protein homolog)-mediated downregulation of MEF2A (myocyte enhancer factor 2A). Circ Heart Fail. 2020;13:e006525.
122. Liu L, Zhang D, Li Y. LncRNAs in cardiac hypertrophy: from basic science to clinical application. J Cell Mol Med. 2020;24:11638-45.
123. Samani NJ, Erdmann J, Hall AS, et al. ; WTCCC and the Cardiogenics Consortium. Genomewide association analysis of coronary artery disease. N Engl J Med. 2007;357:443-53.
124. Rodríguez-Esparragón F, Torres-Mata LB, Cazorla-Rivero SE, et al. Analysis of ANRIL isoforms and key genes in patients with severe coronary artery disease. Int J Mol Sci. 2023;24:16127.
125. Shi P, Tang B, Xie W, et al. LncRNA-induced lysosomal localization of NHE1 promotes increased lysosomal pH in macrophages leading to atherosclerosis. J Biol Chem. 2025;301:110246.
126. Zhu C, Baumgarten N, Wu M, et al. CVD-associated SNPs with regulatory potential reveal novel non-coding disease genes. Hum Genomics. 2023;17:69.
127. Fitzgerald K, White S, Borodovsky A, et al. A highly durable RNAi therapeutic inhibitor of PCSK9. N Engl J Med. 2017;376:41-51.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Special Topic
Copyright

Data & Comments
Data
0











Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].