

Tuning oxygen limitation for polyhydroxyalkanoate production from alkaline lignin liquor fermentation and comparison with nitrogen limitation
 0
0 
Abstract
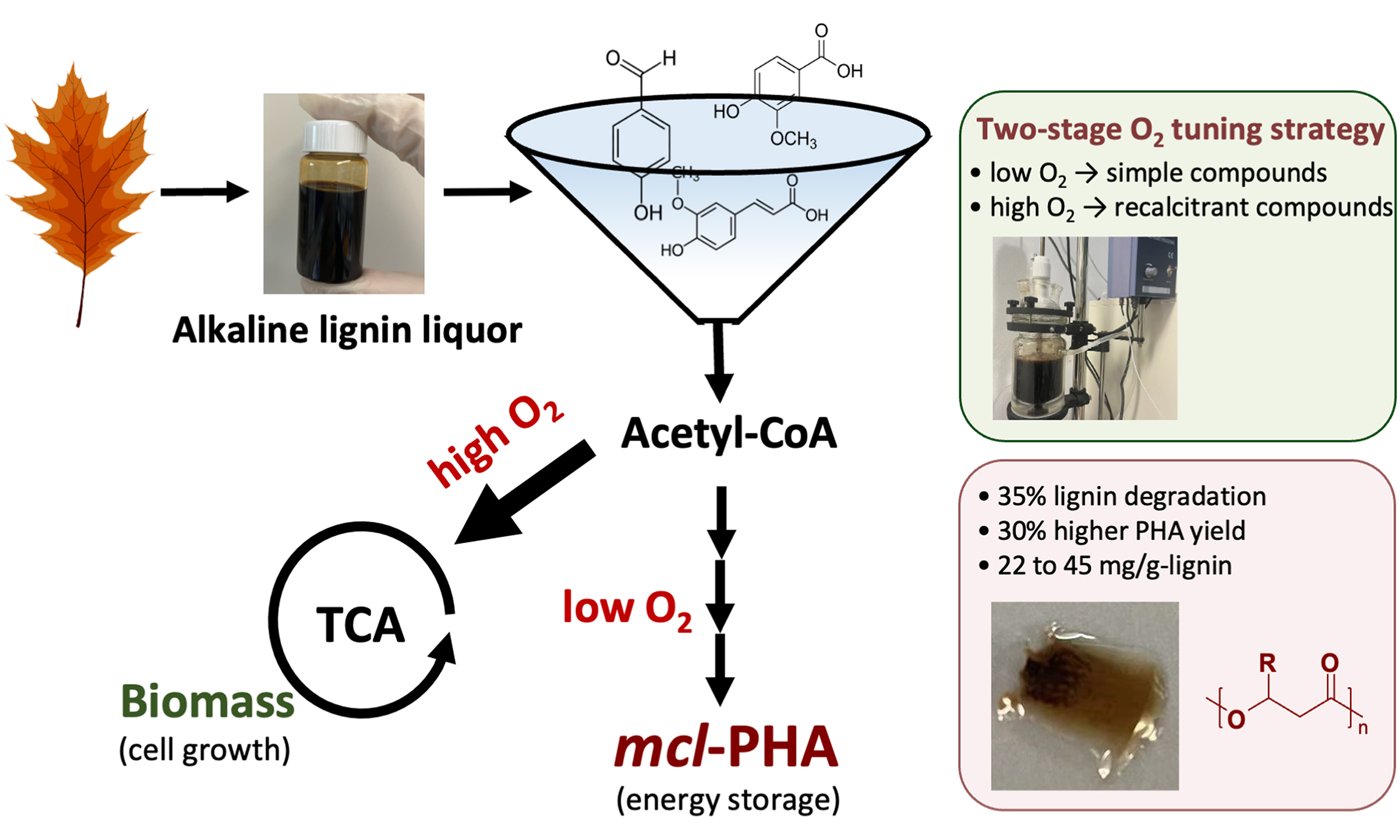
Biosynthesis of lipids/polyhydroxyalkanoates (PHA) enables the valorization of waste into value-added products, yet efficient production has traditionally relied on nutrient limitation to redirect carbon flux. This study proposes an alternative strategy by tuning oxygen limitation during lignin fermentation, targeting lignin as the largest underutilized fraction of lignocellulosic biomass. To address the heterogeneous nature of lignin-derived compounds, a two-stage O2 supplementation strategy was developed: An initial low-oxygen condition (down to 1% dissolved oxygen) facilitated the conversion of readily utilizable compounds, followed by elevated oxygen supply to promote the degradation of more recalcitrant fractions. This strategy yielded PHA of 22 to 45 mg/g-lignin, commensurate with lignin concentrations from 2 to 25 g/L. Notably, significant lignin degradation (comparable to nutrient-replete conditions) and enhanced PHA production (30% higher than nutrient-limited conditions) were simultaneously achieved through staged oxygen supplementation. More broadly, this work demonstrates an alternative to conventional nutrient-limited strategies for regulating the bioconversion of heterogeneous feedstocks into value-added products.
Keywords
INTRODUCTION
Lignocellulose represents vast resources for bio-based chemicals and biofuels, yet lignin remains largely untapped. Modern industrial fermentation is largely centered on carbohydrates - from anaerobic digestion to consolidated bioprocessing and enzymatic saccharification - whereas lignin remains an underutilized carbon resource[1,2]. In the pulp and paper industry, a significant issue is the management of the alkaline lignin liquor from this separation, which amounts to 80 million tons annually, disposed of without valorization[3,4]. Alkali-pretreated lignin can be partially utilized by certain aerobic bacteria, such as Pseudomonas putida (P. putida), to naturally produce polyhydroxyalkanoates (PHA), an energy-rich intracellular polymer that functions as a microbial reservoir of carbon and biochemical energy[5]. This process enables the recovery and storage of lignin-bound energy as biodegradable bioplastics. However, carbon flux diversion limits this process, as substrates are preferentially used for cell growth rather than PHA synthesis during central metabolism, similar to other lipid and PHA biosynthetic pathways[6]. It had been proposed that limiting nutrient supplements, typically nitrogen, could restrict or arrest cell growth while sustaining product synthesis[7,8], which enhanced the biological valorization of glucose, fatty acids, or sole aromatic compounds but found to be less effective for lignin bioconversion[8-10]. This behavior can be attributed to differences in the nature of available carbon sources. Unlike defined single-carbon substrates, lignin-derived compounds are highly heterogeneous and are catabolized hierarchically, with readily utilizable compounds being consumed before more recalcitrant fractions[11,12]. As nitrogen becomes limiting, cells lose the capacity to synthesize new catabolic pathways and extracellular enzymes, thereby restricting the utilization of complex lignin-derived compounds and resulting in low overall lignin degradation (15%-25%)[13].
To date, strategies for enhancing PHA production by redirecting carbon flux have typically relied on restricting cell growth rates (and consequently carbon uptake rates), commonly achieved under nutrient limitation or environmental constraints[8,9,14]. Among these approaches, limiting O2 availability has been used to redirect carbon flux for higher PHA production on catabolizing glucose, glycerol, or fatty acids[14-18]. In the catabolism of aromatics and lignin, as O2 participates in the upper degradation pathways, including enzyme synthesis and acting as an electron acceptor[5,19], oxygen limitation is expected to influence metabolic flux distribution in a manner analogous to nutrient-limitation strategies. Furthermore, given the heterogeneous nature of lignin-derived substrates, where recalcitrant compounds require greater energy input for extracellular enzymatic depolymerization and subsequent utilization[20,21], this study proposes a two-stage oxygen availability strategy. In the first stage, a low-oxygen condition is applied to promote carbon flux toward PHA accumulation while restricting cell growth, allowing the preferential utilization of readily degradable monomeric compounds, similar to conventional nutrient-limited fermentation. In the second stage, oxygen availability is increased to facilitate the bioconversion of more recalcitrant compounds, analogous to nutrient-replete fermentation. With this staged oxygen supplementation strategy, carbon flux is anticipated to be redirected toward enhanced PHA production while maintaining high lignin degradation efficiency. It is noteworthy that Ramírez-Morales et al.[5] have investigated the role of oxygen availability under nitrogen-limited conditions and demonstrated that sufficient oxygen is required to sustain metabolic activity. In contrast, the present study examined the independent effect of oxygen availability under nutrient-unlimited conditions.
In this research, nitrogen-limited lignin fermentation served as the baseline study. Representative monomers present in alkaline lignin liquor and real alkaline lignin liquor were examined for the effect of O2 limitation on their bioconversion. Ultimately, a comparison was conducted between strategies involving O2 limitation and limited nitrogen supplementation. The findings of this study demonstrated the potential of tuning O2 limitation to facilitate lignin bioconversion to PHA, offering an alternative to the conventional nutrient-limited strategy in biological PHA production.
EXPERIMENTAL
Alkaline lignin extraction from yard waste
Yard waste (YW) collected from the campus of the National University of Singapore served as the lignocellulosic feedstock in this study. The material consisted predominantly of fallen leaves (approximately 90% of dry weight) and contained 21% acid-insoluble lignin. After drying, the YW was milled and passed through a 36-mesh sieve prior to pretreatment. To eliminate readily extractable carbohydrates and minimize their carryover into the subsequent lignin stream, the processed YW was first subjected to liquid hot water treatment at a solid loading of
Lignin fermentation
Adjusting O2 limitation
P. putida KT2440 (ATCC 47054) was used in lignin fermentation for PHA production. PHA production was conducted in 250 mL conical flasks using five carbon sources: (1) 20 mM acetate (sodium acetate); (2) 20 mM vanillic acid; (3) 20 mM ferulic acid; (4) 10 g/L lignin. For shake flask experiments, surface aeration was the main source of oxygen supply. To establish varied O2 transfer from air into the culture media during shaking experiments, the volume of media with each carbon source was set to 50, 100, 150, and 200 mL (corresponding to medium depths of 1, 2, 3.5, and 4.8 cm, respectively)[22]. Among these, flasks containing 50 mL of medium represented non-oxygen-limited conditions and were used as the control. Each flask contained 1× modified M9 salts (3 g/L of KH2PO4, 0.5 g/L of NaCl, 6.78 g/L of Na2HPO4) together with 10 mM (NH4)2SO4, 0.2 mM MgSO4, and 10µM CaCl2. The fermentation was conducted at 30 °C with a shaking speed of 225 rpm in a shaking incubator (LM570-RD, YIHDER Tech, China). In experiments where O2 transfer into the medium was further reduced, the fermentation flasks were incubated without shaking, while the culture broth was continuously mixed using a magnetic stirrer, operated at 225 rpm for 72 h at 30 °C. All the experiments were conducted in duplicates.
Lignin fermentation scale-up
The scaled-up lignin fermentation was conducted using a 1 L stirred tank reactor with a working volume of 700 mL. The reactors were maintained at a constant temperature of 30 °C using an outer circulating water bath. Mixing was achieved with a three-blade impeller operating at 225 rpm. Aeration was provided at the bottom of the reactor through varying rates of air sparging, specifically 0.3, 0.1, 0.01, or 0 VVM (Volume of gas per working Volume per Minute). These varying rates were selected to investigate their respective impacts on the fermentation process.
Comparison of tuning O2 limitation and limiting nitrogen strategies
Different concentrations of lignin liquor (2-25 g/L) were fed to P. putida to examine the lignin bioconversion to PHA. In the nitrogen-limited lignin fermentation, the nitrogen source supplement was limited to 1 mM (NH4)2SO4. Other nutrient supplements and environment conditions (e.g., temperature and initial pH) were kept consistent with the conditions described in Adjusting O2 limitation. These conditions were commonly used for cultivation of P. putida. The fermentation was operated in 250 mL shaking flasks with a working volume of
Analytical methods
Lignin quantification
Lignin content was determined following the Klason lignin analytical procedure developed by the National Renewable Energy Laboratory (NREL)[23]. In brief, samples of alkaline lignin liquor were first dried and subsequently hydrolyzed with concentrated sulfuric acid. After centrifugation, the acid-insoluble lignin fraction was quantified gravimetrically, while soluble components remained in the supernatant. To facilitate routine monitoring, the Klason lignin results were correlated with chemical oxygen demand (COD) measurements obtained using a COD reactor system (DBR200, HACH, USA).
Quantification of bacterial growth and polyhydroxyalkanoates
Cell growth and intracellular PHA accumulation were monitored throughout fermentation by measuring optical density at 600 nm (OD600) and Nile Red fluorescence, respectively. To minimize interference from the dark-colored lignin liquor, culture samples were centrifuged at 6,000 × g, washed once with 1× phosphate-buffered saline (PBS), and resuspended in 96-well transparent microplates (Corning Costar, USA) prior to analysis. Measurements were conducted using a microplate reader (Infinite 200 Pro, Tecan, Switzerland). The maximum specific growth rate (μmax) was calculated from the period exhibiting the highest sustained increase in biomass over three consecutive days. For PHA analysis, cell suspensions were stained with Nile Red at a final concentration of 10 μg/mL for 5 min before fluorescence determination. Fluorescence signals were recorded using excitation and emission wavelengths of 485/12 nm and 590/10 nm, respectively. A calibration curve was established between fluorescence intensity and PHA concentrations quantified by gas chromatography (GC), yielding a conversion factor of 113,936 fluorescence units per gram of PHA per liter. The statistical significance levels were determined using a t-test.
Dissolved oxygen measurement
Dissolved oxygen (DO) was measured using a DO meter (Orion Star A213 RDO/DO, ThermoFisher). Prior to measurement, the DO meter was calibrated using a two-point calibration (zero calibration and air-saturated water calibration). Briefly, Milli-Q water sparged with nitrogen gas for 1 h, followed by adding 2% Na2SO3, was used for zero calibration while Milli-Q water sparged with air for 1 h was used for 100% air-saturated water calibration.
Measurement of molecular weight of lignin
The molecular weight distribution of lignin was quantified using gel filtration chromatography (GFC) to characterize water-soluble compounds. Briefly, 2 g/L of raw or fractionated alkaline lignin liquor was filtrated (0.22 μm syringe filters) and then subjected to GFC analysis (Agilent 1260 infinity ii) with refractive index detector. The eluent was 0.02% sodium azide and the flow rate was 1.0 mL/min. An injection volume of 30 μL was used.
RESULTS AND DISCUSSION
Baseline of nitrogen-limited lignin fermentation
To establish a baseline of lignin fermentation with nitrogen-limited strategy, different concentrations of alkaline lignin liquor (2-25 g/L) were used as the substrates for examination. The relationship between cell growth and lignin concentration is shown in Figure 1A. Increasing lignin concentration resulted in monotonic increase of cell growth within 24 h except the group at 25 g/L feedstock, in which peak biomass was only reached at 48 h, caused by the inhibition posed by high concentration lignin on microbial activity. The groups with 2-10 g/L feedstock sustained cell viability while biomass decreased with lignin concentrations exceeded 15 g/L, indicating nitrogen starvation. Lignin degradation extent decreased with increasing lignin concentrations, achieving maximum and minimum degradation extents of 29% and 16% when fermenting with 2 g/L and 25 g/L lignin, respectively [Figure 1B]. It is noteworthy that this degradation is lower than that under nitrogen-replete conditions (35% degradation)[20,24]. Lignin utilization increased monotonically with feeding concentration, even under nitrogen limitation [Supplementary Figure 1], as higher lignin levels supplied more readily available carbon substrates that were preferentially consumed, consistent with hierarchical substrate utilization[11,12]. This further indicates that more recalcitrant compounds are not effectively catabolized under growth-restricted conditions. In terms of PHA production, the lowest values recorded at 25 g/L lignin with yield of
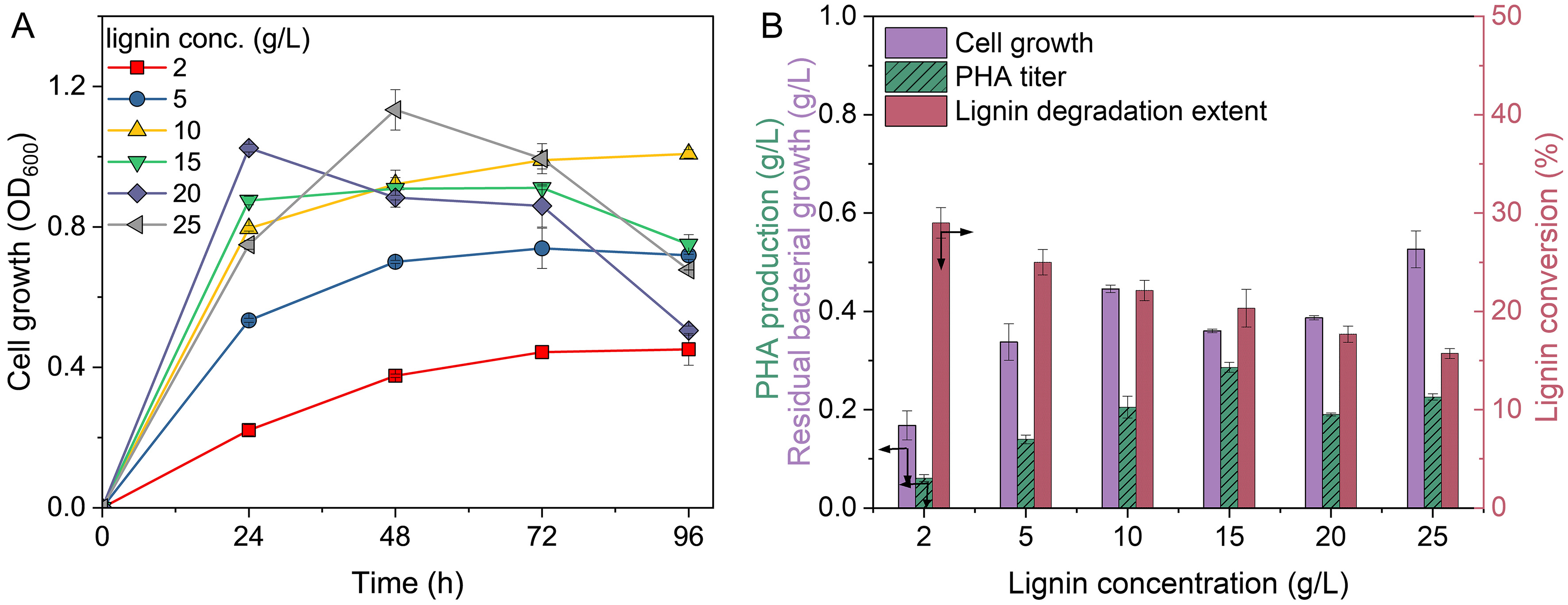
Figure 1. (A) Cell growth and (B) lignin bioconversion at different feeding concentrations (2-25 g/L) using nitrogen-limited strategy. Error bars represent the standard deviation of two replicates (n = 2). OD600: Optical density at 600 nm; lignin conc.: lignin concentration; PHA: polyhydroxyalkanoates.
Effects of O2 limitation on degradation of lignin monomers
Microbes exhibit varying growth condition requirements when metabolizing different carbon sources[11], necessitating an understanding of the requisite O2 limitation for converting individual lignin compounds. As monomeric compounds in alkaline lignin liquor comprise linear carboxylic acids and aromatic compounds, three monomers identified in alkaline lignin liquor[21] - acetate, vanillic acid, and ferulic acid - were selected as the representative substrates to investigate the effect of O2 limitation on lignin bioconversion. Different volumes of media (ranging from 50 to 200 mL) were used in the shaking flasks to establish varying levels of O2 limitation, as the rate of oxygen diffusion into the medium decreases with increasing volume (surface aeration)[27]. The 50 mL condition was used as the positive control, representing non-oxygen-limited conditions. Figure 2 presents the effect of limiting O2 availability on cell growth (OD600). To characterize the impacts of O2 limitation on redirecting carbon fluxes (ratio of PHA to cell growth), the resulting residual bacterial biomass (dcw excluding PHA concentration) and PHA production are shown in Figure 3. The results presented in Figures 2A and 3 indicate that O2 limitation did not restrict the growth (maximum specific growth rate μmax of 0.03OD600/h) of P. putida on acetate, resulting in similar carbon flux redirection with a PHA concentration of 19% PHA/dcw across all the flasks. Although acetate was slightly volatile, limiting O2 supplementation reduced substrate loss, marginally improving terminal bacterial growth from 0.3 g/L in 50 mL medium to 0.4 g/L in 150 mL medium, concomitant with PHA production of about 0.1 g/L. This unrestricted cell growth on acetate by varying O2 availability was attributed to its simple degradation pathway, which could directly enter central carbon metabolism via acetyl-CoA [Supplementary Figure 2]. Acetate was measured at the end of fermentation, and no residual carbon source was detected (data not shown).
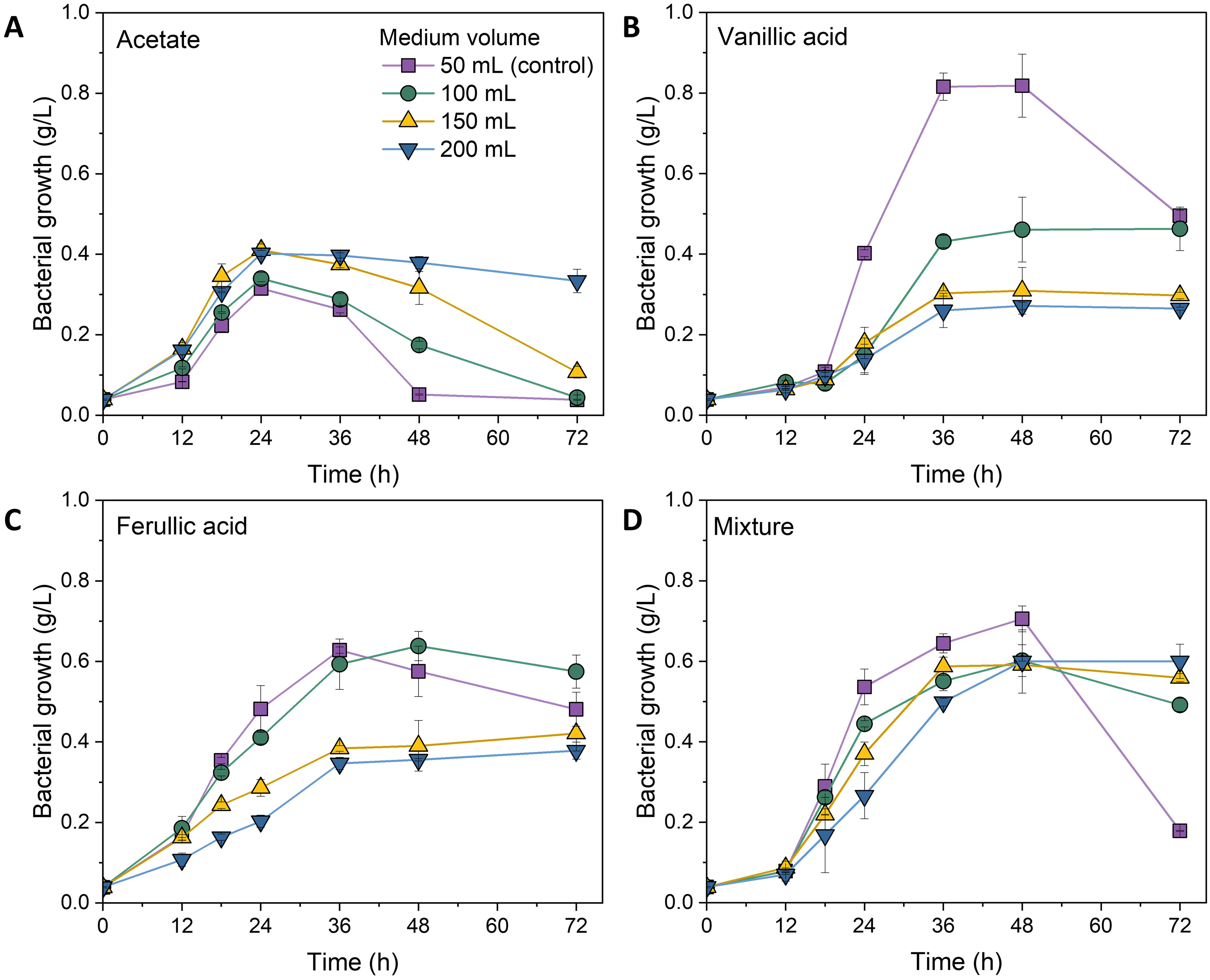
Figure 2. Bacterial growth (OD600) of P. putida under O2-limited conditions on the sole carbon source of (A) acetate, (B) vanillic acid, (C) ferulic acid, and (D) their mixture at 6.67 mM each. Error bars represent the standard deviation of two replicates (n = 2). OD600: Optical density at 600 nm; P. putida: Pseudomonas putida.
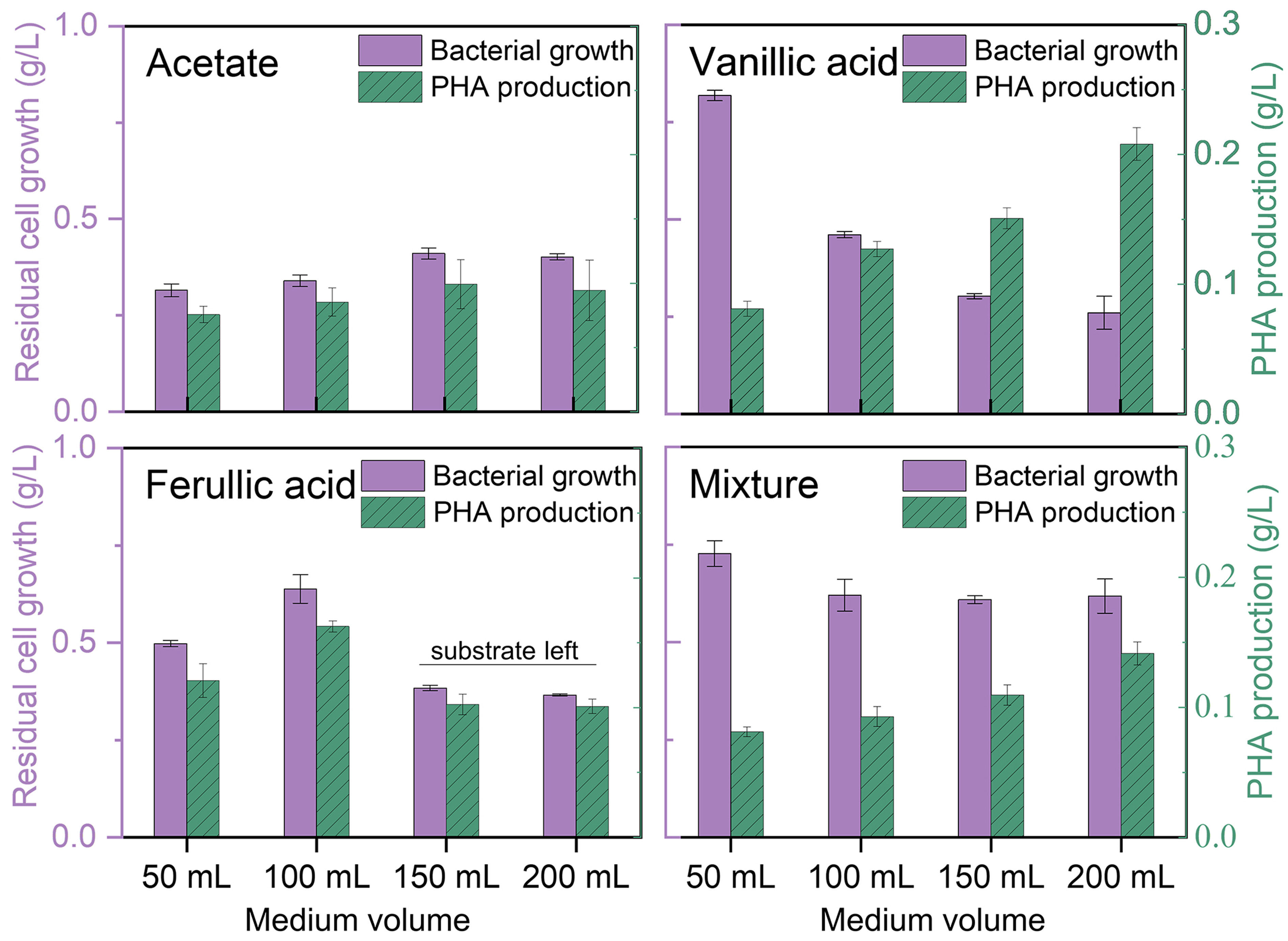
Figure 3. Peak PHA production and its corresponding cell growth of P. putida within 72 h on acetate, vanillic acid, ferulic acid as the sole carbon source, and their mixture at 6.67 mM each under O2-limited conditions. Error bars represent the standard deviation of two replicates (n = 2). PHA: Polyhydroxyalkanoates; P. putida: Pseudomonas putida.
Aromatic compounds undergo upper degradation to protocatechuic acid, followed by aromatic ring opening via the β-ketoadipate pathway to central carbon metabolism[19]. Molecular oxygen is not only necessary for energy metabolism; the catabolism of vanillic acid and ferulic acid involves multiple oxygen-dependent enzymatic cleavages and oxidation reactions prior to entering central carbon metabolism [Supplementary Figure 2]. When utilizing vanillic acid as the sole carbon source, increasing O2 limitation reduced the cell growth rate with μmax of 0.06, 0.03, and 0.01OD600/h in 50, 100, and
The catabolism of ferulic acid, a more recalcitrant aromatic monomer than vanillic acid, involves initial enzymatic cleavages to acetyl-CoA and vanillin. Acetyl-CoA is directly utilized in central carbon metabolism, while vanillin is further degraded following the same catabolic pathway as vanillic acid [Supplementary Figure 2]. Ferulic acid degradation can also occur extracellularly by enzymes secreted via outer membrane vesicles, leading to the appearance of acetate and vanillic acid in the extracellular environment[28,29]. Considering that acetyl-CoA derived from ferulic acid degradation theoretically supports a maximum cell growth of about 0.6 OD600 (acetate utilization in Figure 2A), the varied cell growth on ferulic acid within the first 24 h - from the highest at 0.74 OD600 in 50 mL flasks to the lowest at 0.31 OD600 in 200 mL flasks [Figure 2C] - demonstrates that the cleavage of ferulic acid to acetyl-CoA and vanillic acid was affected by O2 limitation. As the appearance of the easily utilizable carbon source acetyl-CoA, which could accelerate the utilization of aromatic compounds, increasing O2 limitation by increasing medium volume from 50 to 100 mL did not significantly restrict cell growth (P > 0.05), resulting in similar carbon flux redirection with PHA content of about 20% PHA / dcw [Figure 2C and Figure 3]. The marginal increase in cell growth and PHA production in the 100 mL flask compared to the 50 mL flask was attributed to reduced acetate volatilization. Further increases of O2 limitation in the 150 and 200 mL flasks reduced both cell growth and PHA production due to substrate leftovers. At the end of the 72-h fermentation, intermediate vanillic acid from ferulic acid catabolism was detected in the 150 mL and
Alkaline lignin comprises a collection of linear and aromatic compounds. Considering that P. putida’s response to O2 limitation might differ between utilizing sole and mixed substrates, three selected monomeric compounds - acetate, vanillic acid, and ferulic acid - were mixed at equivalent molar concentrations (6.67 mM each) to examine the effect of O2 limitation on the bioconversion of mixed lignin monomers. As shown in Figure 2D and Figure 3, increasing O2 limitation resulted in a monotonic decrease in cell growth while increasing PHA production, with PHA content (PHA/dcw) rising from 10% in 50 mL flasks to 18% in 200 mL flasks. This demonstrated the redirection of P. putida carbon fluxes towards higher PHA production on mixed substrates through O2 limitation. Additionally, it is important to note that P. putida’s cell growth on mixed substrates was enhanced compared to that on individual sole carbon sources. Under the highest level of O2 limitation (200 mL medium), fermentation using mixed substrates doubled the maximum specific growth rate (μmax of 0.027 OD600/h) compared to that on sole vanillic acid (μmax of 0.013) or ferulic acid (μmax of 0.015) [Figure 2]. This demonstrates that the simple carbon source acetate facilitated the utilization of the aromatic compounds. This result suggested that fermentation on mixed substrates such as alkaline lignin liquor might require further increasing O2 limitation to restrict cell growth and reconfigure more fluxes towards PHA production.
Tuning O2 limitation on bioconversion of alkaline lignin liquor
Appropriate O2 limitation has been confirmed to restrict cell growth on aromatic monomers and enhance PHA production. In this section, the monomers were replaced by 10 g/L alkaline lignin liquor to examine the effects of O2 limitation on lignin bioconversion. Similar to the examination of lignin monomer bioconversion, varying O2 limitation was achieved by using different volumes of medium (50-200 mL) in 250 mL shaking flasks. With the same level of O2 limitation as in the fermentation of lignin monomers, the results of residual cell growth, PHA production, and lignin degradation in Figure 4A-C show that O2 limitation during lignin fermentation did not exhibit a clear trend in either cell growth or PHA production, resulting in a similar final PHA production across all the flasks, at approximately 0.2 g/L. This insignificant carbon flux redirection was attributed to an unrestricted cell growth rate. As shown in Figure 4A, cell growth in all the flasks increased rapidly to above OD600 of 1.0 within 24 h, reaching over 80% of the total cell biomass in 120 h. This trend was consistent with the findings from fermentation on mixed monomers [Figure 2D], where the cell growth on mixed substrates was enhanced compared to that on the sole carbon sources. Alkaline lignin liquor represents a more complex mixture of substrates, consisting of many linear compounds like acetate and malonic acid, as well as aromatics[21]. During fermentation of alkaline lignin liquor, the utilization of linear compounds could facilitate the degradation of aromatics, explaining the insignificant restriction of cell growth at the same level of O2 limitation used for restricting cell growth on aromatic monomers.
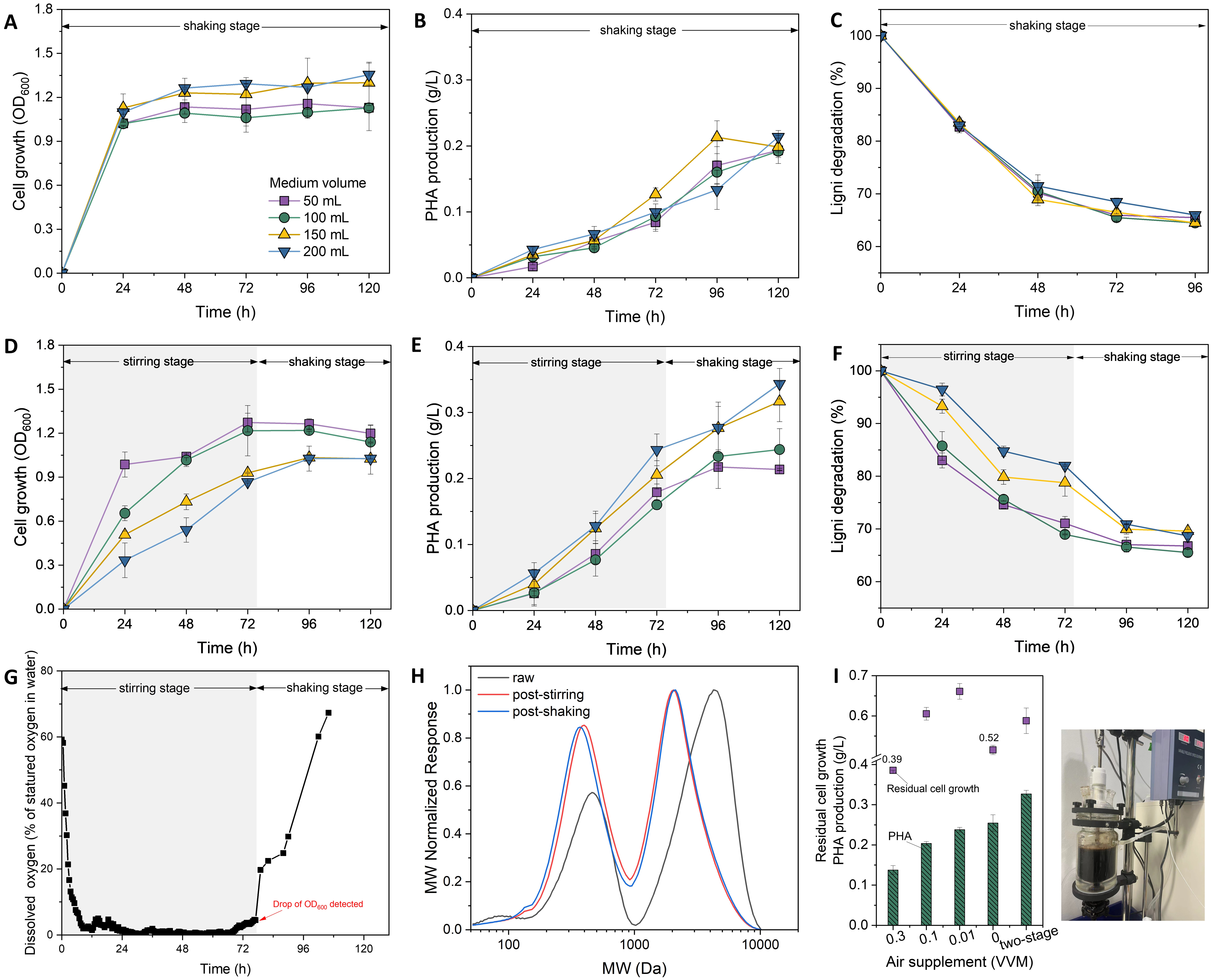
Figure 4. (A) Residual cell growth, (B) PHA production, and (C) lignin degradation during lignin bioconversion in shaking flasks with different volumes of medium. Residual cell growth represents the increase of dcw excluding PHA; (D) Residual cell growth, (E) PHA production, and (F) lignin degradation during lignin bioconversion using stirring followed by shaking flasks with different volumes of medium; (G) Dissolved oxygen in flasks with 150 mL medium using the approach of stirring followed by shaking flask; (H) Molecular weight distribution of lignin liquor from raw feedstock, post-stirring, and post-shaking fermentation; (I) Scaled-up lignin bioconversion in a 1 L reactor with different levels of air supplementation. Photograph of the experimental setup taken by the authors. Error bars represent the standard deviation of two replicates (n = 2). PHA: Polyhydroxyalkanoates; MW: molecular weight.
To further restrict cell growth on alkaline lignin, O2 limitation was intensified by performing lignin fermentation in flasks with only medium stirring rather than flask shaking. This approach of only stirring the medium effectively distinguished the maximum specific cell growth rate and resulting bacterial biomass (within the first 72 h) [Figure 4D]. Maximum PHA production within 72 h reached 0.24 g/L [Figure 4E], surpassing the total PHA produced by fermentation in shaking flasks. After 72 h of fermentation in stirring flasks, substrate utilization tended to stagnate [Figure 4F]; therefore, the fermentation was transferred into a shaking incubator for shaking flasks, thereby increasing oxygen transfer into the medium and facilitating the degradation of the most recalcitrant compounds. At the end of the shaking stage, flasks containing 50 and 100 mL of medium showed a similarly high level of cell growth at OD600 of 1.2. Flasks with 150 and 200 mL of medium restricted cell growth to about OD600 of 1.0 and enhanced the terminal PHA production to 0.31 and 0.34 g/L, respectively, approximately 45% higher than in 50 and 100 mL of medium [Figure 4E].
Examination of DO levels facilitated understanding of the relationship between fermentation outcomes and O2 availability. Figure 4G illustrates the DO levels in the 150 mL flasks. During the stirring stage, DO decreased to below 1% of the saturation level in water, during which cell growth and PHA synthesis could be maintained, as shown in Figure 4D and E. This was consistent with the observation that P. putida could sustain PHA synthesis at 0% DO when utilizing simple carbon sources[18]. When the stirring stage proceeded to 72 h, lignin degradation stagnated, and DO rose, guiding the decision to change the oxygen supply approach to using shaking flasks, where DO gradually rose above 20% and finally reached above the level of 80%. Molecular weight distribution analysis identified two dominant peaks in the lignin liquor [Figure 4H], corresponding to low-molecular-weight compounds (< 1,000 Da) and a higher-molecular-weight fraction centered at approximately 5,000 Da. P. putida was able to utilize compounds from both fractions. The molecular weight profile of the abiotic aeration control remained unchanged relative to the raw lignin feedstock, indicating that lignin degradation was primarily attributable to microbial activity rather than aeration-induced oxidation [Supplementary Figure 4].
Lignin fermentation in 1 L stirred tank reactor
Industrial fermentation typically employs impellers and air spargers for agitation and oxygen supplementation. To validate this approach for industrial applications, lignin fermentation was scaled up in a 1 L tank reactor equipped with a three-blade impeller and bottom air sparger. Different levels of air supplementation ranging from 0.3 to 0 VVM were examined [Figure 4I and Supplementary Figure 5]. Results indicated that reducing air supplementation from 0.3 to 0.01 VVM monotonically increased both cell growth and PHA production, achieving PHA of 0.13, 0.20, and 0.24 g/L with air supply at 0.3, 0.1, and 0.01 VVM, respectively. This was probably caused by substrate volatilization and reduced cell viability at high oxygen supply[30], as reflected by the lack of corresponding increases in cell growth and PHA despite reduced lignin levels [Supplementary Figure 4]. The aeration-induced substrate loss was further confirmed through a separate abiotic experiment evaluating substrate loss under different aeration conditions [Supplementary Figure 6]. Further reduction of air supply from 0.01 to
Comparison with nitrogen-limited strategy
As traditional approaches to enhance PHA production often involved redirecting carbon flux by limiting nutrient supplementation, particularly nitrogen[8,9], it was appropriate to compare the approaches of tuning O2 limitation with nitrogen limitation on lignin bioconversion. For this comparison, different concentrations (2-25 g/L) of lignin were examined to compare the results from nitrogen-limited fermentation in Figure 1. Figure 5 and Supplementary Figure 7 present the results of lignin bioconversion, cell growth, and PHA production using a two-stage tuning O2 limitation approach during the stirring stage (0-72 h) and shaking stage (0-144 h), respectively. At the end of the stirring stage, fermentation with low lignin concentrations (2 and 5 g/L) had achieved their maximum degradation of 37% and 30%, respectively, which were not significantly enhanced during the shaking stage (P > 0.05). Lignin degradation decreased to 18% with
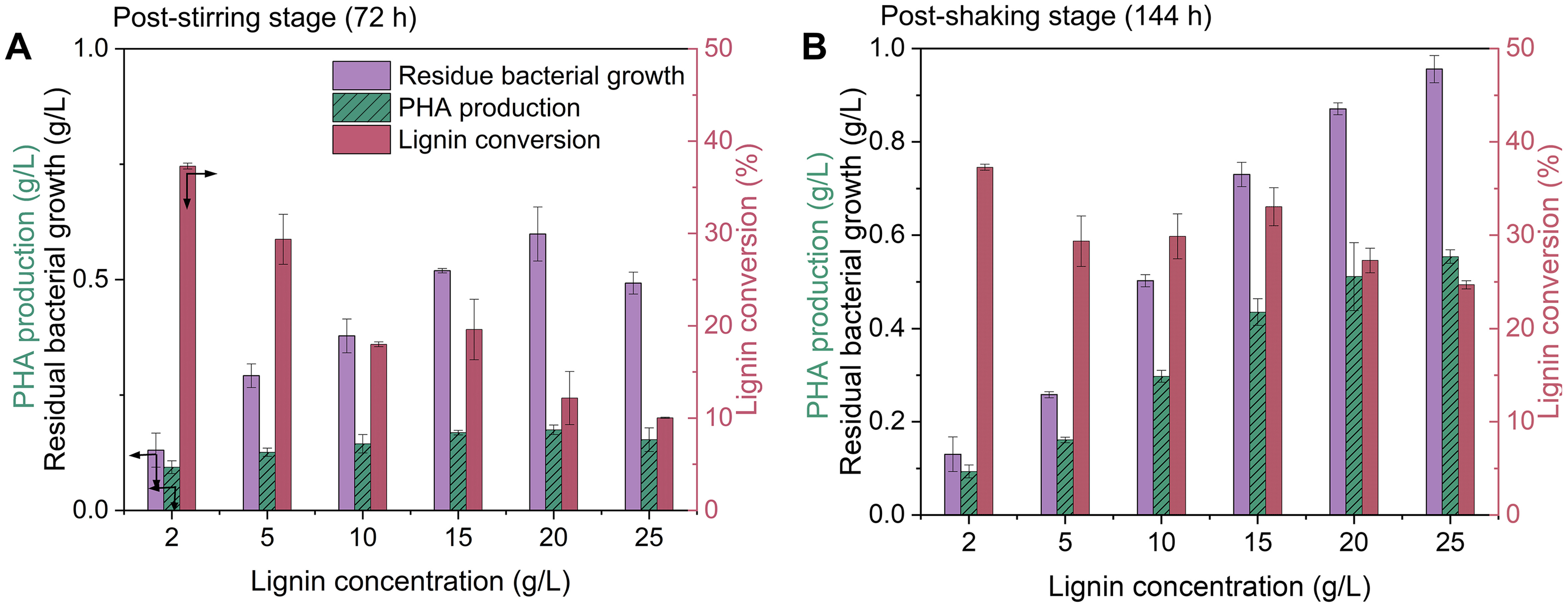
Figure 5. Lignin bioconversion at different feeding concentrations through tuning O2 limitation in flasks (A) after the stirring stage and (B) after the shaking stage. Error bars represent the standard deviation of two replicates (n = 2). PHA: Polyhydroxyalkanoates.
Comparing the results of lignin bioconversion under these two strategies, the approach of tuning O2 limitation demonstrated several advantages over the nitrogen-limited strategy in redirecting carbon fluxes towards PHA production, achieving higher lignin degradation, and enhancing treatment capacity for high lignin concentrations. Firstly, O2 limitation during the stirring stage imposed a stronger restriction on cell growth, resulting in increased PHA production compared to the nitrogen-limited strategy. Evidence from fermentations with 2 and
PHA compositions
Understanding PHA composition across different strategies is crucial for determining bioplastic properties and potential applications. For instance, short-chain-length PHA (C4-C6) exhibits brittleness, high melting temperature, crystallinity, and tensile strength, making it suitable for packaging, whereas medium-chain-length PHA (C8-C10) is flexible and finds applications in adhesives and biomedical applications[31,32]. In this study, when O2 was limited, 3-hydroxydecanoic acid (C10) accounted for over 80% of the PHA composition with lignin at 2 and
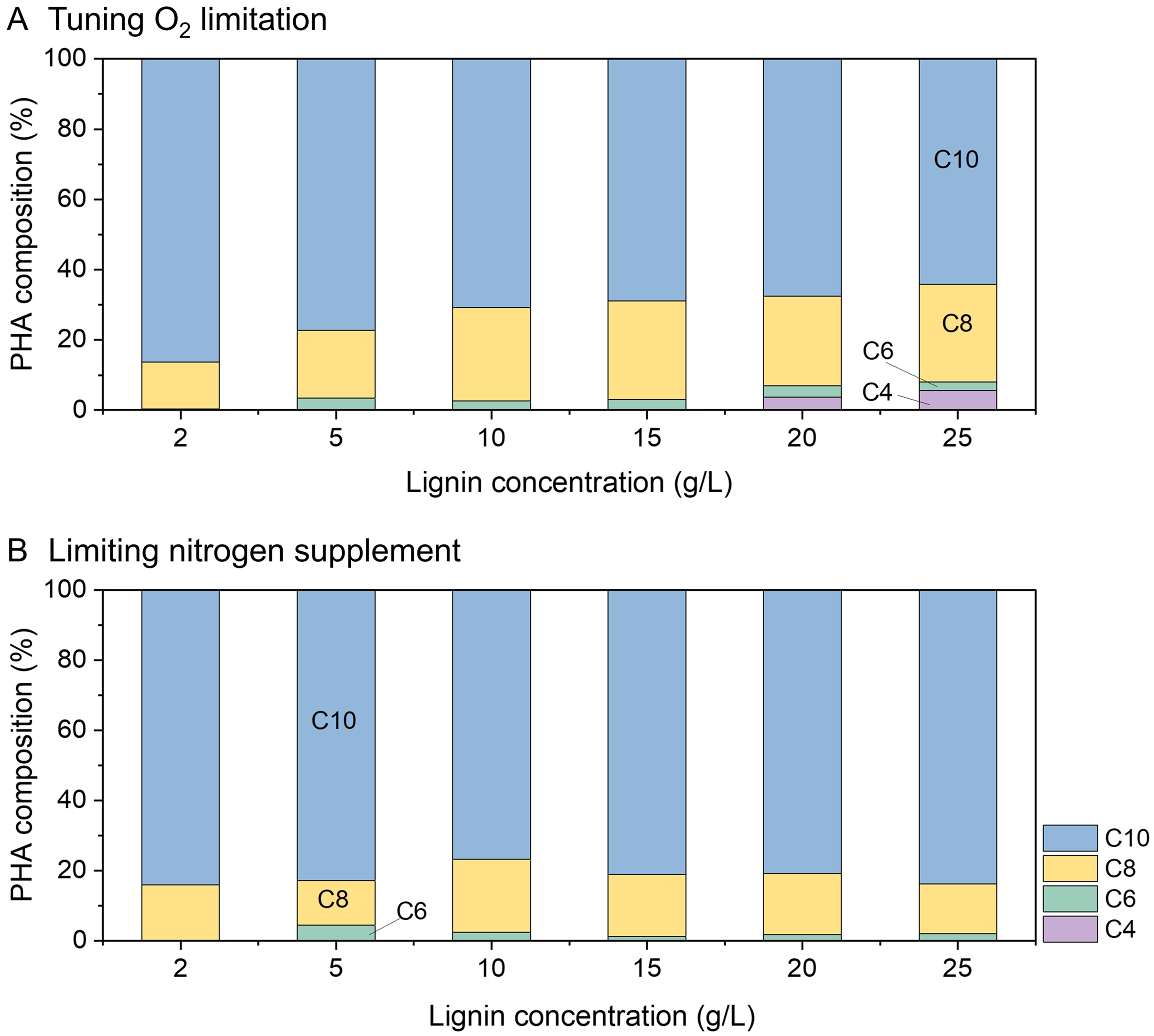
Figure 6. PHA composition in fermentation under varying lignin concentrations using strategies involving (A) tuning O2 limitation and (B) nitrogen limitation. C4-C10 represent hydroxy fatty acid monomers obtained from the hydrolysis of the produced PHA. PHA: Polyhydroxyalkanoates.
Discussion and perspectives
Limiting nitrogen supply enables rapid cell growth to a moderate density, at which cells retain the capacity to catabolize the same carbon source for PHA production. However, due to the heterogeneous nature of lignin-derived substrates and their hierarchical utilization, nitrogen depletion restricts the catabolism of more complex compounds[12]. To explore an alternative approach, this study examined the effect of oxygen availability on the fermentation of compounds in alkaline lignin liquor. During the low-oxygen stage (initial 72 h; Figures 4 and 5), carbon source utilization was reduced, while PHA accumulation increased. This phenomenon has also been observed for other substrates, such as glucose and butanol, although the underlying genetic mechanisms remain unclear[16,17]. In the subsequent stage (72-144 h; Figures 4 and 5), where oxygen was no longer limiting, fermentation proceeded under non-limited conditions, enabling the utilization of more recalcitrant compounds and further enhancement of PHA production. Notably, 1 L tank reactor experiments demonstrated that lignin fermentation can be sustained under surface aeration without additional air sparging, and that reducing aeration rates helps minimize substrate loss [Figure 4I and Supplementary Figure 5], which is critical for future industrial applications. Further work is recommended to elucidate the mechanisms of metabolic flux redistribution, particularly the decoupling between reduced carbon utilization rates and enhanced PHA accumulation. A techno-economic analysis is warranted to evaluate industrial-scale implementation. Nutrient-limited fermentation benefits from reduced nutrient consumption but may experience carbon losses during aeration, whereas the oxygen-tuning strategy enables higher lignin conversion. The trade-off between nutrient savings and improved carbon utilization should be assessed to determine the most economically viable approach.
CONCLUSIONS
This study has demonstrated the feasibility of manipulating O2 limitation to modulate metabolic fluxes during lignin bioconversion. It highlighted the benefits of low O2 availability in directing more easily utilizable compounds towards PHA, while improved O2 availability enhanced the bioconversion of recalcitrant compounds. The approach of tuning O2 limitation had proven effective in processing high concentrations of lignin liquor, resulting in PHA yields ranging from 22 to 45 mg/g-lignin, depending on lignin feeding concentrations of 2 to 25 g/L. This strategy is particularly advantageous for the bioconversion of heterogeneous substrates, offering an alternative to conventional nutrient-limited strategies in biological PHA production.
DECLARATIONS
Authors’ contributions
Conceptualization, methodology, investigation, data curation, writing - original draft: Sun, J.
Writing - review and editing, funding acquisition: Tong, Y. W.
Writing - review and editing, supervision, funding acquisition: Loh, K. C.
Availability of data and materials
The data supporting the findings of this study are presented in this manuscript and Supplementary Materials.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
This research project was funded by the National Research Foundation, Prime Minister’s Office, Singapore under its Campus for Research Excellence and Technological Enterprise (CREATE) Program.
Conflicts of interest
Tong, Y. W. is an Associate Editor of the Advanced Energy Conversion journal. He had no involvement in the review or editorial process of this manuscript, including but not limited to reviewer selection, evaluation, or the final decision, while the other authors have declared that they have no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
Supplementary Materials
REFERENCES
1. Sun, J.; Loh, K. C. Navigating the interface between lignocellulose pretreatment and bioconversion in lignin-inclusive fermentation: a review. Bioresour. Technol. 2026, 439, 133325.
2. Liu, Z. H.; Hao, N.; Wang, Y. Y.; et al. Transforming biorefinery designs with ‘Plug-In Processes of Lignin’ to enable economic waste valorization. Nat. Commun. 2021, 12, 3912.
3. Chen, X.; Mosier, N.; Ladisch, M. Valorization of lignin from aqueous-based lignocellulosic biorefineries. Trends. Biotechnol. 2024, 42, 1348-62.
4. Mboowa, D. A review of the traditional pulping methods and the recent improvements in the pulping processes. Biomass. Conv. Bioref. 2021, 14, 1-12.
5. Ramírez-morales, J. E.; Czichowski, P.; Besirlioglu, V.; et al. Lignin aromatics to PHA polymers: nitrogen and oxygen are the key factors for Pseudomonas. ACS. Sustain. Chem. Eng. 2021, 9, 10579-90.
6. Nikel, P. I.; Fuhrer, T.; Chavarría, M.; Sánchez-pascuala, A.; Sauer, U.; De Lorenzo, V. Reconfiguration of metabolic fluxes in Pseudomonas putida as a response to sub-lethal oxidative stress. ISME. J. 2021, 15, 1751-66.
7. Cezairliyan, B.; Ausubel, F. M. Investment in secreted enzymes during nutrient-limited growth is utility dependent. Proc. Natl. Acad. Sci. U.S.A. 2017, 114, E7796-E7802.
8. Elmore, J. R.; Dexter, G. N.; Salvachúa, D.; et al. Production of itaconic acid from alkali pretreated lignin by dynamic two stage bioconversion. Nat. Commun. 2021, 12, 2261.
9. Kacanski, M.; Stelzer, F.; Walsh, M.; Kenny, S.; O’connor, K.; Neureiter, M. Pilot-scale production of mcl-PHA by Pseudomonas citronellolis using acetic acid as the sole carbon source. New. Biotechnol. 2023, 78, 68-75.
10. Ling, C.; Peabody, G. L.; Salvachúa, D.; et al. Muconic acid production from glucose and xylose in Pseudomonas putida via evolution and metabolic engineering. Nat. Commun. 2022, 13, 4925.
11. Mendonca, C. M.; Yoshitake, S.; Wei, H.; et al. Hierarchical routing in carbon metabolism favors iron-scavenging strategy in iron-deficient soil Pseudomonas species. Proc. Natl. Acad. Sci. U.S.A. 2020, 117, 32358-69.
12. Sun, J.; Loh, K. One-Pot lignin bioconversion to polyhydroxyalkanoates based on hierarchical utilization of heterogeneous compounds. Bioresour. Technol. 2025, 419, 132056.
13. Salvachúa, D.; Karp, E. M.; Nimlos, C. T.; Vardon, D. R.; Beckham, G. T. Towards lignin consolidated bioprocessing: simultaneous lignin depolymerization and product generation by bacteria. Green. Chem. 2015, 17, 4951-67.
14. Yen, H. W.; Zhang, Z. Effects of dissolved oxygen level on cell growth and total lipid accumulation in the cultivation of Rhodotorula glutinis. J. Biosci. Bioeng. 2011, 112, 71-4.
15. Zhang, X.; Chen, J.; Wu, D.; Li, J.; Tyagi, R. D.; Surampalli, R. Y. Economical lipid production from Trichosporon oleaginosus via dissolved oxygen adjustment and crude glycerol addition. Bioresour. Technol. 2019, 273, 288-96.
16. Ankenbauer, A.; Nitschel, R.; Teleki, A.; et al. Micro‐aerobic production of isobutanol with engineered Pseudomonas putida. Eng. Life. Sci. 2021, 21, 475-88.
17. Lu, H.; Chen, H.; Tang, X.; et al. Metabolomics analysis reveals the role of oxygen control in the nitrogen limitation induced lipid accumulation in Mortierella alpina. J. Biotechnol. 2021, 325, 325-33.
18. Blunt, W.; Dartiailh, C.; Sparling, R.; Gapes, D.; Levin, D. B.; Cicek, N. Microaerophilic environments improve the productivity of medium chain length polyhydroxyalkanoate biosynthesis from fatty acids in Pseudomonas putida LS46. Process. Biochem. 2017, 59, 18-25.
19. Jiménez, J. I.; Miñambres, B.; García, J. L.; Díaz, E. Genomic analysis of the aromatic catabolic pathways from Pseudomonas putida KT2440. Environ. Microbiol. 2002, 4, 824-41.
20. Kruger, J. S.; Dreiling, R. J.; Wilcox, D. G.; et al. Lignin alkaline oxidation using reversibly-soluble bases. Green. Chem. 2022, 24, 8733-41.
21. Linger, J. G.; Vardon, D. R.; Guarnieri, M. T.; et al. Lignin valorization through integrated biological funneling and chemical catalysis. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 12013-8.
22. Somerville, G. A.; Proctor, R. A. Cultivation conditions and the diffusion of oxygen into culture media: the rationale for the flask-to-medium ratio in microbiology. BMC. Microbiol. 2013, 13, 9.
23. Sluiter, A.; Hames, B.; Ruiz, R.; et al. Determination of Structural Carbohydrates and Lignin in Biomass. NREL/TP-510-42618. Laboratory Analytical Procedure (LAP). National Renewable Energy Laboratory, 2008. https://www.researchgate.net/publication/285273316_Determination_of_structural_carbohydrates_and_lignin_in_biomass_in_Laboratory_Analytical_Procedure_LAP#read (accessed 2026-06-24).
24. Lin, L.; Cheng, Y.; Pu, Y.; et al. Systems biology-guided biodesign of consolidated lignin conversion. Green. Chem. 2016, 18, 5536-47.
25. Arreola-vargas, J.; Xu, B.; Hu, C.; et al. Bioconversion of agave bagasse lignin to medium-chain-length polyhydroxyalkanoates by Pseudomonas putida. ACS. Sustain. Chem. Eng. 2022, 10, 15670-9.
26. Arreola-vargas, J.; Meng, X.; Wang, Y.; Ragauskas, A. J.; Yuan, J. S. Enhanced medium chain length-polyhydroxyalkanoate production by co-fermentation of lignin and holocellulose hydrolysates. Green. Chem. 2021, 23, 8226-37.
27. Maier, U.; Losen, M.; Büchs, J. Advances in understanding and modeling the gas-liquid mass transfer in shake flasks. Biochem. Eng. J. 2004, 17, 155-67.
28. Choi, C. W.; Park, E. C.; Yun, S. H.; et al. Proteomic characterization of the outer membrane vesicle of Pseudomonas putida KT2440. J. Proteome. Res. 2014, 13, 4298-309.
29. Salvachúa, D.; Werner, A. Z.; Pardo, I.; et al. Outer membrane vesicles catabolize lignin-derived aromatic compounds in Pseudomonas putida KT2440. Proc. Natl. Acad. Sci. U.S.A. 2020, 117, 9302-10.
30. Meilhoc, E.; Wittrup, K. D.; Bailey, J. E. Influence of dissolved oxygen concentration on growth, mitochondrial function and antibody production of hybridoma cells in batch culture. Bioprocess. Eng. 1990, 5, 263-74.
31. Muthuraj, R.; Valerio, O.; Mekonnen, T. H. Recent developments in short- and medium-chain- length Polyhydroxyalkanoates: production, properties, and applications. Int. J. Biol. Macromol. 2021, 187, 422-40.
32. Rai, R.; Keshavarz, T.; Roether, J.; Boccaccini, A.; Roy, I. Medium chain length polyhydroxyalkanoates, promising new biomedical materials for the future. Mater. Sci. Eng. R. Rep. 2011, 72, 29-47.
33. Cha, D.; Ha, H. S.; Lee, S. K. Metabolic engineering of Pseudomonas putida for the production of various types of short-chain-length polyhydroxyalkanoates from levulinic acid. Bioresour. Technol. 2020, 309, 123332.
34. Manoli, M.; Blanco, F. G.; Rivero-buceta, V.; et al. Heterologous constitutive production of short-chain-length polyhydroxyalkanoates in Pseudomonas putida KT2440: the involvement of IbpA inclusion body protein. Front. Bioeng. Biotechnol. 2023, 11, 1275036.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0









Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at [email protected].